12 maggio 2008 - 12:17 pm
Forse questo post avrebbe dovuto essere tra i primissimi, però meglio tardi che mai. Mi ricordo di aver letto su un testo di patologia questa definizione: un virus è una brutta notizia avvolta in una proteina. In effetti quando si parla di virus si parla sempre di brutte – o pessime – notizie: sono organismi talmente semplici da essere sulla linea di confine tra la materia vivente e la materia non vivente, talmente poveri strutturalmente da aver bisogno dell’aiuto di una cellula ospite per riprodursi, sono perciò patogeni obbligati. In quanto tali, non stupisce che causino malattia, a volte lieve, a volte molto grave: ecco il perché della brutta notizia. La seconda parte della definizione fa riferimento alla struttura comune a tutti i virus: uno strato proteico che ricopre il genoma. Molti virus si limitano a questa struttura, altri aggiungono strati aggiuntivi, quasi sempre formati da fosfolipidi. Andiamo con ordine e cominciamo con la storia.
Storia della virologia
La prima volta che si è cercato di contrastare un’infezione virale è stata quando, nel 1796, Edward Jenner cominciò la sua campagna di vacinazione contro il vaiolo. Dell’esperimento ne abbiamo già parlato (nel post sui vaccini), c’è però da dire che allora non si aveva idea di cosa fosse un virus. Questo fu un primo tentativo empirico, che per fortuna risultò enormemente valido. Tanto che Pasteur, un secolo dopo, chiamò la sua procedura vaccinazione, in onore agli esperimenti di Jenner. Egli coniò anche il termine Virus, prendendo la parola dal latino (veleno). Da quel momento ci fu un’impennata di conoscenza sui virus: si isolarono i corpi inclusi dalle infezioni di vaiolo, successivamente si scoprì che esisteva un agente infettivo filtrabile, più piccolo di qualunque batterio conosciuto: il virus del mosaico del tabacco (TMV).
Nel 1900 venne dimostrato che la febbre gialla era trasmessa dalle zanzare e, nel 1911, Rous dimostrò che alcuni virus causavano tumore. Si diede un senso al detto “anche le pulci hanno le pulci” quando, nel 1915, si scoprì che alcuni virus possono infettare dei batteri. Questa scoperta diede uno slancio enorme nella ricerca in virologia, perché dava l’opportunità di lavorare praticamente senza rischi.
Nel 1940 finalmente la prima foto: veniva fotografato il TMV al microscopio elettronico, e nel 1952 Harshey e Chase dimostrarono, con l’uso di un batteriofago, che il codice genetico risiede nel DNA e non nelle proteine.
Oggi le conoscenze sono moltissime, la struttura dei virus, la loro biologia molecolare e la loro biochimica sono conosciuti molto bene, ma è ovvio che queste prime scoperte furono fondamentali, come è anche ovvio che le conoscenze di oggi sono ancora limitate e migliorabili. La storia della virologia è costellata di grandi scoperte (alcune da premio Nobel) e da grandi abbagli (come quando si credeva che il kuru, malattia prionica, fosse data da un virus). Ma questo credo sia comune un po’ a tutta la biologia.
Struttura
Esistono virus di tutti i tipi: piccoli o molto grandi, allungati, a forma di mattone o proiettile, rotondi, icosaedrici… Le dimensioni variano da 18 a 700 nm di diametro (anche se esistono i filovirus, con particella allungata, la cui lunghezza può arrivare ad 1 μm) e contengono un solo tipo di acido nucleico: esistono infatti virus a DNA e virus a RNA.
Il capside

Il genoma dei virus è di norma costituito da una catena di acido nucleico, che può essere DNA o RNA, a seconda del tipo di virus, ed è normalmente contenuto in una struttura proteica chiamata capside. Il capside proteico può avere diverse forme: elicoidale, icosaedrico, complesso (come nei fagi T pari o il capside a mattone del Poxvirus). Quando il virus è nudo, il capside è l’unico rivestimento della particella. In questo caso presenta le proteine necessarie per il riconoscimento recettoriale indispensabile per l’infezione (spikes o spicole).
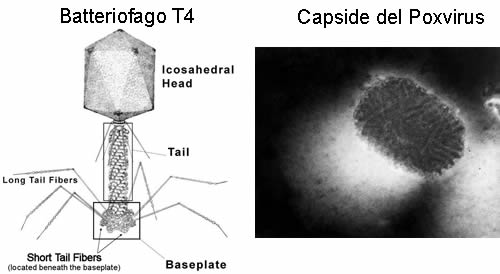
Molti virus hanno invece un secondo rivestimento, proveniente dalla membrana cellulare della cellula ospite, e formato da fosfolipidi. Questo secondo rivestimento è chiamato envelope, su cui sono evidenti le spikes necessarie per l’infezione, che non fanno parte del capside ma che sono vere e proprie proteine di membrana, spesso glicoproteine. Esistono poi altri tipi di proteine necessarie all’infezione, si va dalle proteine importanti per la fusione delle membrane a quelle emoagglutinanti, l’importante è capire che comunque tutte sono indispensabili al virus: in una struttura piccola ed essenziale come quella virale non c’è spazio per strutture inefficenti. Nei virus con envelope, il capside e il genoma virale sono spesso chiamati con l’unico nome di nucleocapside, mentre il capside è spesso chiamato core.
Esistono poi, nei virus con envelope, delle proteine con funzione strutturale, situate proprio al di sotto dello strato fosfolipidico, chiamate proteine di matrice o proteine M.
Infine, alcuni virus portano con se degli enzimi: HIV ha al suo interno la trascrittasi inversa e l’integrasi, ma nei virus possono trovarcisi anche polimerasi, o complessi importanti per la maturazione dell’RNA. Nessuno di loro però ha ribosomi, o meccanismi metabolici per produrre ATP, quindi nessuno di loro è autosufficiente, sono quindi parassiti obbligati.
Tags:
capside,
core,
envelope,
spicole,
virus nudo
10 febbraio 2008 - 11:35 am
 Replicazione
Replicazione
I diversi ceppi di virus influenzale sono tutti capaci di replicarsi nella cavità amniotica di uova di pollo embrionate , che in laboratorio rappresenta il modo migliore per la propagazione del virus; inoltre molti ceppi si moltiplicano bene in diversi tipi di colture cellulari e nei tratti respiratori di furetti e topi.
L’ingresso del virus nella cellula avviene tramite l’interazione tra la proteina HA e l’acido sialico: questo comporta l’internalizzazione del virus tramite endosoma. La fuoriuscita del virus dalla vescicola avviene dopo l’acidificazione, che causa un cambio conformazionale nell’HA, causando l’esposizione del peptide fusogeno che fonde l’envelope virale e la mambrana dell’endosoma, liberando nel citoplasma il core. A differenza di altri virus a RNA, in Orthomyxovirus la replicazione è nucleare.
PB1, PB2 e PA hanno insieme la funzione di RNA-polimerasi RNA-dipendente (RNA-trascrittasi), e trascrivono il filamento di mRNA a partire dagli RNA a filamento negativo del genoma virale. Il virus utilizza meccanismi eucariotici per aggiungere il cap e la catena di poliA, per questo motivo ha necessità di entrare nel nucleo della cellula.
L’RNA messaggero trasloca nel citoplasma dove è tradotto, si formano le proteine del capside (NP) e gli enzimi PB1, PB2 e PA che saranno presenti nei virioni maturi, inoltre vengono trascritte copie dell’RNA a filamento negativo che sarà il corredo genomico del virus. La maturazione di HA e NA avvene a livello del Golgi, dove vengono glicosilate, per poi essere trasportate alla membrana cellulare. A questo punto avviene l’assemblaggio del virus: anche il nucleocapside è trasportato in membrana e il virus esce per gemmazione. La proteina NA esplica a questo livello il suo ruolo: taglia i residui di acido sialico in modo da evitare interfereze tra la membrana e i virus appena formati.
Patologia
A livello cellulare il virus può causare principalmente due effetti: inibizione della sintesi delle proteine cellulari (con annessa aumento della sintesi di quelle virali) e stress ossidativo.
Il primo effetto si ha sia sulla sintesi di nuove proteine, sia sul blocco della traduzione di proteine che stanno già nascendo. Il virus agisce sul complesso proteico CBP (Cap binding protein): esso è formato da tre subuntuà: eIF4E, eIF4A ed eIF4G, che insieme formano il CBP, o eIF4F. Avviene una fosforilazione della subunità 4A e una defosforilazione di 4E, come conseguenza abbiamo una aumentata efficienza di traduzione per gli mRNA virali e un’inibizione di quelli cellulari.
Il secondo effetto è dato dalla comparsa, durante l’infezione virale, di specie reattive dell’ossigeno.
A livello clinico la malattia si presenta come una polmonite, e i sintomi sono conosciuti da tutti, tanto da essere presi a confronto con quelli di altre malattie: febbre, dolori diffusi, stanchezza, inappetenza, che però non sono causati dal virus in sé, che raramente da viremie, ma da prodotti dell’infezione, come frammenti di cellule morte o citochine di origine leucocitaria. Normalmente è autolimitante, le situazioni più gravi si trovano in pazienti già debilitati oppure in presenza di sovrainfezioni batteriche.
Tags:
core,
emoagglutinina,
envelope,
influenza,
neuraminidasi,
orthomyxovirus,
RNA-trascrittasi
 Commenti disabilitati
Commenti disabilitati
8 febbraio 2008 - 11:04 am
 La fine della prima guerra mondiale coincise con una catastrofe sanitaria che portò più vittime della stessa guerra: 20 milioni di morti in meno di un anno. Tra la fine del 1918 e l’inizio del 1919 l’influenza colpì con una violenza mai vista, espandendosi rapidamente in tutto il mondo e facendo il doppio delle vittime rispetto alla guerra appena finita. Questa epidemia fu chiamata “la spagnola”, non perché fosse originata in Spagna, ma perché la Spagna era neutrale e non era sottoposta a censure sui giornali, così diede per prima la notizia. Non fu certo la prima, e nemmeno l’ultima tra le gravi epidemie di influenza, ma di sicuro fu quella che causò più danni.
La fine della prima guerra mondiale coincise con una catastrofe sanitaria che portò più vittime della stessa guerra: 20 milioni di morti in meno di un anno. Tra la fine del 1918 e l’inizio del 1919 l’influenza colpì con una violenza mai vista, espandendosi rapidamente in tutto il mondo e facendo il doppio delle vittime rispetto alla guerra appena finita. Questa epidemia fu chiamata “la spagnola”, non perché fosse originata in Spagna, ma perché la Spagna era neutrale e non era sottoposta a censure sui giornali, così diede per prima la notizia. Non fu certo la prima, e nemmeno l’ultima tra le gravi epidemie di influenza, ma di sicuro fu quella che causò più danni.
Struttura e classificazione
Il virus influenzale (Orthomyxovirus) è l’agente eziologico della classica influenza stagionale, che si presenta tipicamente come una infezione delle vie aeree superiori o con una polmonite. Sono stati isolati tre tipi di virus: A, B e C, l’ultimo dei quali molto raramente causa epidemie, inoltre esistono moltissimi sottotipi, alcuni umani, altri aviari, altri ancora che infettano altri mammiferi.
La forma e le dimensioni del virus possono essere vari, ma generalmente si presentano come sferici o ovoidali, con un diametro di circa 100 nm (qui delle foto). Il virione si presenta rivestito da un envelope lipidico su cui troviamo varie proteine strutturali (le proteine di matrice o M), non strutturali (NS) e due glicoproteine transmembrana: emoagglutinina (HA) e neuraminidasi (NA), importanti per l’adsorbimento e l’ingresso del virus nella cellula.
All’interno dell’envelope troviamo il core virale, dove è contenuto il genoma, formato da 8 molecole di RNA a singolo filamento a polarità negativa, nucleoproteine del capside (NP) associate ad essi, e tre proteine importanti per la replicazione del genoma: PB1, PB2 e PA.
Antigenicamente si distinguono delle varianti all’interno dei gruppi A e B, le quali però non sono associate tanto alla traduzione delle proteine, quanto agli zuccheri associati alle glicoproteine. Inoltre però esistono, all’interno del gruppo A, ulteriori differenziazioni che fanno riferimento alla struttura primaria delle due proteine HA e NA, dando così origine a vari gruppi di virus quali H1N1 (responsabile della spagnola), H2N2, H5N1 e così via.
Il virus influenzale può cambiare antigenicamente tramite due meccanismi: antigenic drift e antigenic shift. Il primo tipo di variazione causa i cosiddetti cambiamenti antigenici minori, e si basa sul meccanismo classico delle mutazioni puntiformi del genoma. Il secondo invece è detto anche cambiamento antigenico maggiore, ed è causato da un fenomeno di ricombinazione.
Antigenic shift. La ricombinazione virale può esistere solo nel caso in cui due virus diversi infettino la stessa cellula: nell’assemblamento dei virioni alcuni frammenti di RNA del primo virus si mischieranno con altri del secondo, dando origine così ad un nuovo virus, con caratteristiche diverse da quelli originali. Nel 1919 questo fenomeno ha portato alla pandemia che ha ucciso 20 milioni di persone: un virus umano e uno aviario si incontrarono in un ospite che poteva essere infettato da entrambi: il maiale. Dopo il fenomeno di antigenic shift avvenuto nel maiale, il virus acquistò la capacità di infettare esseri umani e di trasmettersi tra loro, senza bisogno di passaggi intermedi in altri animali.
Questa è la paura odierna nei riguardi dell’H5N1: il virus, normalmente aviario, ha acquistato la capacità di infettare anche gli uomini, ma non di propagarsi tra loro. La malattia portata dall’H5N1 è più grave dell’influenza classica, ma è di difficile diffusione, se dovesse incrociarsi con un ceppo umano ci potrebbero essere molti problemi nel contenerla.
Fonti:
R. Dulbecco, H.S. Ginsberg: Virologia
Tags:
antigenic shift,
core,
emoagglutinina,
envelope,
influenza,
neuraminidasi,
orthomyxovirus
Commenti disabilitati
