
 L’insonnia è una delle malattie più diffuse del nostro tempo: chi non ha passato almeno una notte insonne, per ritrovarsi il mattino dopo incapace di ricordare con facilità nozioni o concetti? Per non parlare di come diventa difficile mantenere la concentrazione e portare avanti una conversazione di senso compiuto. Negli scorsi post vi ho raccontato di come l’insonnia alteri la memoria, oggi vi parlo di come il dormire male porti problemi a comunicare. Nelle api, e forse anche nell’uomo.
L’insonnia è una delle malattie più diffuse del nostro tempo: chi non ha passato almeno una notte insonne, per ritrovarsi il mattino dopo incapace di ricordare con facilità nozioni o concetti? Per non parlare di come diventa difficile mantenere la concentrazione e portare avanti una conversazione di senso compiuto. Negli scorsi post vi ho raccontato di come l’insonnia alteri la memoria, oggi vi parlo di come il dormire male porti problemi a comunicare. Nelle api, e forse anche nell’uomo.
Uno sguardo ai meccanismi della mente
Inside Neuroscience
8 giugno 2011 - 5:07 pm
Il sonno e la comunicazione: la danza delle api

28 aprile 2011 - 5:09 pm
Il Coma (III parte): Il risveglio
Preambolo
Rieccoci per continuare di nuovo il nostro viaggio nell’affascinante mondo delle neuroscienze. Nei post precedenti abbiamo introdotto alcuni concetti del coma con delle considerazioni basilari (1, 2), questa volta, invece, ci focalizzeremo sul “risveglio” e dei possibili trattamenti.
Però, prima di ri-entrare nel merito dell’argomento, sono necessarie alcune considerazioni ovvie. Questo non è un blog medico come neanche l’autore di questo testo, quindi non c’è alcuna intenzione di fuorviare dalle terapie affidate da medici competenti, né tantomeno di procurare delle informazioni pericolose per automedicazioni improvvisate. Ogni riferimento di questo testo dovrebbe essere preso per quello che è, ovvero un ampliamento delle conoscenze di base, un pretesto per destare curiosità nell’ambito specifico, e dare un minimo di erudizione.
Cenni di anatomia
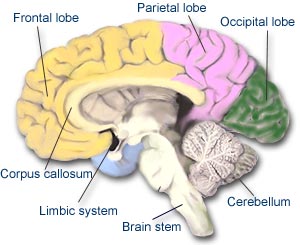
Di fatti, ‘svegliare’ qualcuno dal coma significa liberare o accendere il proprio ‘io’, ma se andate a cercare su un libro di anatomia, fisiologia, o neuroscienze dove si trovi effettivamente il nostro ‘io’ resterete delusi, per ora è ancora un mistero. La biologia della coscienza è uno dei processi mentali più complessi, e di conseguenza è anche il meno compreso. La nostra personalità ed il nostro stato di attenzione è deciso da un insieme di neuroni localizzati nel profondo del nostro cervello ed è difficile determinare come questi interagiscano per dare vita ad una unica personalità.
Grossolanamente, dividiamo il nostro encefalo in tre parti: il cervello, il cervelletto ed iltronco encefalico.
- Il cervello rappresenta l’85% del peso totale dell’encefalo ed è la parte più complicata del nostro organismo; controlla l’intelligenza, la ragione, la memoria, le emozioni, la visione ed i sentimenti, etc. Sicuramente detiene almeno una parte cospicua della nostra coscienza.
- Il cervelletto è la parte più piccola dell’encefalo. Risiede alla base della nuca e gioca un ruolo essenziale nella coordinazione, la postura ed il bilanciamento dell’organismo.
- Il tronco encefalico è la parte del nostro encefalo che connette il cervello con il midollo spinale ed è responsabile per il controllo di molte funzioni basilari come il respiro, pressione sanguigna, lo stato di veglia e l’attenzione.
Curiosamente, anche se non sappiamo esattamente dove sia il nostro ‘io’ all’interno del cervello, sappiamo che una parte del tronco encefalico è il suo ‘interruttore’. Numerose evidenze, infatti, hanno dimostrato che se vogliamo ‘accendere’ o ‘liberare’ il nostro ‘io’ dobbiamo interferire con una particolare regione del tronco encefalico, ovvero il romboecefalo. Le neuroconnessioni tra questa piccola regione del midollo allungato e la corteccia determinano lo stato di veglia, il sonno o lo stato comatoso.
Tuttavia queste informazioni sembrano quasi inutili se paragonate alla complessità del coma stesso.
Perché occuparsi del coma?
Tutti i vari stati di incoscienza sono sintomi di disfunzioni neurologiche severe. Infatti, é buona norma preoccuparsi seriamente ed indagare a fondo se osserviamo una persona che perde conoscenza improvvisamente e senza un alcun motivo apparente. Si potrebbe trattare di narcolessia, epilessia, stati di shock emotivi o patologie meno gravi del coma, ma in ogni caso possono comunque avere degli effetti indiretti drammatici durante la guida, il lavoro o semplicemente mentre si attraversa la strada.
Tra queste patologie sicuramente il coma é tra le più invalidanti sebbene possa essere una condizione transitoria.
Tra le più diffuse patologie che possono causare il coma in modo silente sono da ricordare il diabete mellito, la pressione sanguigna alta, i problemi renali, i disturbi al fegato oppure gli attacchi epilettici. Inoltre, sono da considerare anche l’abuso di alcool, traumi cerebrali, overdose da stupefacenti ed addirittura la carenza di zuccheri.
I sintomi premonitori
Esistono dei sintomi premonitori per il coma, ma purtroppo hanno delle caratteristiche molto diverse, ad esempio il coma può iniziare immediatamente senza alcun sintomo evidente, o si può sviluppare lentamente nel tempo con caratteristiche varie. Generalmente la progressione dello stato di incoscienza e come velocemente si sviluppa sono già dei possibili indizi su cosa l’abbia causato.
Altre variabili che si tengono in conto sono il ritmo del respiro, le funzioni cardiovascolari, le caratteristiche della pelle, problemi evidenti agli arti, i riflessi oculari ed altre cose di questo genere. A volte, specifiche anormalità possono essere utili per identificare il problema cerebrale scatenante. Purtroppo però, nella maggioranza dei casi non si hanno informazioni conclusive, per cui sono necessarie delle indagini strumentali molto approfondite.
Come si può curare?
Sicuramente il trattamento del coma dipende da caso a caso, tuttavia se la causa è sconosciuta, i medici possono dare al paziente un cosiddetto “cocktail per il coma” che può essere utile in alcuni casi di emergenza. Questa terapia preliminare non è fatta di altro che da un insieme di vitamine, zuccheri ed alcuni farmaci; ovviamente non é una terapia efficace e tanto meno universale. Può essere utile per risolvere alcuni casi meno gravi degli stadi comatosi. Le vitamine aiutano i pazienti con problemi nutrizionali o di abuso di alcool, così come gli zuccheri che aiutano i pazienti con un calo di glucosio. La funzione delle altre sostanze presenti nel cocktail é quella di revertire l’azione dei narcotici più diffusi. Ovviamente in una seconda fase d’emergenza, bisogna indagare più a fondo per capire quale sia la causa del coma e trattarlo nel modo più opportuno possibile.
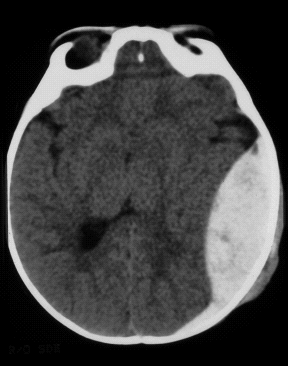
Il caso più intuibile é il coma indotto da un trauma cerebrale. Il rigonfiamento per edema che ne consegue incrementa la pressione intracranica e rallenta il flusso sanguigno in alcune aree con conseguente perdita della coscienza. Una variazione sul tema è un danneggiamento del cervello che causa lo spostamento di alcune aree cerebrali esercitando una pressione sui tessuti circostanti, comprimendo i vasi sanguigni locali. Questo evento particolare, chiamato ‘erniazione cerebrale’, può portare al coma ed in seguito alla morte se non trattato immediatamente.
Ovviamente il risveglio da questo tipo di coma potrebbe facilmente verificarsi spontaneamente in seguito all’abbassarsi della pressione intracranica che si può ottenere per riassorbimento spontaneo, terapia osmotica o chirurgica. La probabilità di recupero dipende ovviamente, dalle zone cerebrali interessante, dalla gravità del danno e dalla prontezza ad iniziare la terapia giusta. In genere questi casi traumatici possono facilmente portare a morte il paziente o ad un rapido recupero, tuttavia, ci possono essere delle rare eccezioni, come nel caso del sig. Donald Herbert, un pompiere andato in coma dopo essere rimasto sepolto sotto un crollo, che dopo 10 anni si risveglia spontaneamente e ritorna in piena attività. Probabilmente è stato il trauma cerebrale a causare il coma, ma é ancora un mistero cosa lo abbia effettivamente risvegliato dopo tutto questo tempo.
Purtroppo sono pochissime le storie con questo lieto fine. Ne é un esempio la sig.ra Christa Smith, andata in coma in seguito ad un attacco cardiaco per poi ‘migliorare’ lentamente fino ad entrare in stato vegetativo. La speranza che questa donna di 50 anni possa migliorare ancora è tanta, perché si è svegliata più volte dallo stato vegetativo ed altrettante volte è ritornata in stato di incoscienza. I parenti stanno ancora con le dita incrociate ed attendono, giorno dopo giorno con il fiato sospeso, qualche novità dall’ospedale.
Coma da intossicazione o infezione
Per il coma da intossicazione, invece, si possono avere dei sintomi che si sviluppano lentamente, da una leggera confusione con sintomi di sonnolenza, e/o ci possono anche essere dei repentini cambi di personalità fino al vero e proprio coma. Purtroppo, ancora oggi non si é chiaro cosa determini precisamente lo stato di coma e quindi non é noto neanche quale sia la procedura migliore per il ‘risveglio’.
Consideriamo tali pazienti come intrappolati nella propria mente, magari da un corto circuito neuronale o da una mancata neurotrasmissione. La cosa interessante é che in molti casi di ‘risvegli’ sono stati somministrati dei farmaci che interferiscono proprio con le neurotrasmissioni centrali. É stato il caso di Amy Pickard che entrò in coma per una overdose di eroina e che poi é stata risvegliata per puro caso dopo la somministrazione di un farmaco ipnotico che altera transitoriamente le neurotrasmissioni del SNC. Il suo coma è durato circa 6 anni e sarebbe potuto durare anche di più se questo farmaco non avesse causato un’interferenza tale da interrompe o alterare il corto circuito neuronale che provocava il coma.
Forse anche per questo motivo in passato si é provato a dare dei cocktail di farmaci per il sistema nervoso centrale con la speranza di riuscire ad alterare lo stato comatoso e causare un risveglio. Purtroppo questi risultati, insieme allo shock elettrico, non hanno mai fornito dei risultati incoraggianti. Quello di Amy é stato un rarissimo caso positivo su un milione di tentativi. Forse é stata usata casualmente la dose o il momento giusto per la somministrazione, oppure il farmaco corretto per quel tipo di ‘blocco’ cerebrale. Più probabilmete era una combinazione di tutti questi fattori ed il particolare stato comatoso della paziente. Di certo non si é mai verificato un altro risveglio dal coma con lo stesso farmaco.
Casi eclatanti

Altri casi eclatanti sono stati Jesse Ramirez e Terry Willis che erano stati dati per spacciati dai medici poiché si trovavano stabilmente in coma da molti anni. In entrambi i casi si pensò di staccare il sondino gastrico per la nutrizione e da lì a pochi giorni Jesse Ramirez uscì dall’ospedale camminando da solo in pieno possesso delle proprie facoltà mentali, mentre Terry Willis è stato ‘meno fortunato’ poiché si è ‘semplicemente’ svegliato dopo 20 anni di coma, e per uscire dall’ospedale ha dovuto usare una sedia a rotelle.
Purtroppo devo sottolineare ancora una volta che i casi citati si possono contare con le dita delle mani. Nella quasi totalità dei casi non c’é la pagina finale del libro, e talvolta non si considerano le sofferenze fisiche dei pazienti in coma per decenni e le sofferenze psicologiche dei parenti. La frase ‘…e vissero tutti felici e contenti’ é sostituita da una lunga via crucis di piaghe, trattamenti, attese snervanti, ed in definitiva da una intrinseca sofferenza che si stenta a descrivere. Forse potremmo impropriamente racchiudere tutto questo nella frase simbolica ‘…a volte penso che sarebbe meglio se…’.
A tal proposito spesso si dimenticano anche altri tipi di storie, come il caso di Sarah Scantlin che in seguito ad un terribile incidente resta in uno stato vegetativo da incubo. Era in grado solo di battere le palpebre, e secondo le analisi il suo cervello era severamente danneggiato da non poter capire quello che succedeva intorno. Ebbene questa donna si ‘risveglia’ dopo 20 anni di coma/stato vegetativo e comincia a parlare, ricorda tantissimi fatti accaduti mentre era in stato di ‘incoscienza’ descrivendo anche la sua terribile sofferenza. Ovviamente non ha recuperato tutte le sue facoltà mentali, poiché i danni cerebrali erano troppo vasti, tuttavia è ancora in grado di ragionare, parlare, muoversi e sta lentamente riprendendosi la sua vita ‘interrotta’ 20 anni prima.
Sono i casi come questi che fanno ri-pensare seriamente al significato di ‘stato di incoscienza’ del coma o dello stato vegetativo e ricordano ancora una volta che il cervello non é del tutto ‘spento’ e perso durante il coma. Questo può dare speranza ad alcuni ed in altri può dare un certo grado di inquietudine per la condizione umana dell’ammalato.
Conclusione
Sicuramente il cervello è un organo estremamente complesso ed ogni suo ‘malfunzionamento’ può essere solo notato; é rarissima la possibilità di comprendere tali fenomeni, ancor di meno correggerli. Spesso ci troviamo come davanti ad un complesso ‘sistema televisivo’ che in qualche caso dà segnali di malfunzionamento. A volte basta un pugno nel punto giusto o una sequenza di tasti per poter riportare il sistema in funzione, tuttavia nella maggioranza dei casi il ‘guasto’ richiede delle risoluzioni più raffinate che ancora oggi non riusciamo a trovare.
Alla prossima
Tags: cervello, coma, risveglio, trauma cerebrale1 marzo 2011 - 8:04 pm
Ho perso il sonno e…la memoria!
 Dormire poco può far male...Vi sarà sicuramente capitato di passare una notte insonne. E, il giorno dopo, concentrarsi e riuscire a portare a termine le attività quotidiane è davvero molto difficile. Il sonno è necessario,anzi, fondamentale ma resta tutt’ora un illustre sconosciuto. Perchè si dorme? Cosa accade nel cervello durante questo stato di incoscienza? E cosa succede se, al contrario, non dormiamo abbastanza?Negli scorsi post vi ho accennato a come il sonno sia coinvolto con il consolidamento della memoria. Che dire se poi si scopre che questo meccanismo ha un ruolo addirittura nello svilupparsi di “false memorie”? Continue Reading »
Dormire poco può far male...Vi sarà sicuramente capitato di passare una notte insonne. E, il giorno dopo, concentrarsi e riuscire a portare a termine le attività quotidiane è davvero molto difficile. Il sonno è necessario,anzi, fondamentale ma resta tutt’ora un illustre sconosciuto. Perchè si dorme? Cosa accade nel cervello durante questo stato di incoscienza? E cosa succede se, al contrario, non dormiamo abbastanza?Negli scorsi post vi ho accennato a come il sonno sia coinvolto con il consolidamento della memoria. Che dire se poi si scopre che questo meccanismo ha un ruolo addirittura nello svilupparsi di “false memorie”? Continue Reading »
9 novembre 2010 - 7:01 pm
Oligodendrociti e mielina. Se non è solo questione di materia grigia…
 …potrebbe essere colpa della “materia bianca”!
…potrebbe essere colpa della “materia bianca”!
Non è solo la corteccia cerebrale, la cosiddetta “materia grigia”, ad essere importante per le funzioni cognitive: sono sempre di più le evidenze sperimentali che mostrano come abbiano grande influenza la mielina, gli oligodendrociti e la “materia bianca” in senso più esteso. Continue Reading »
Tags: apprendimento, ATP, depressione, encefalo, materia bianca, mielina, Neuroni, oligodendrociti, risonanza magnetica, schizofrenia, sonno22 ottobre 2010 - 7:55 pm
Memoria e apprendimento: chi ha spostato il mio formaggio?
 Non si smette mai d’imparare… Già, ma come si fa? O meglio, cosa accade nel nostro cervello quando siamo impegnati ad apprendere e memorizzare un’informazione?
Non si smette mai d’imparare… Già, ma come si fa? O meglio, cosa accade nel nostro cervello quando siamo impegnati ad apprendere e memorizzare un’informazione?
Un recente studio pubblicato su Nature Neuroscience fa il punto della situazione, dirigendo i riflettori su quello che viene definito “apprendimento spaziale”.
Stiamo parlando di quanto succede quando ad esempio dovete ricordarvi la posizione di un oggetto nello spazio. Continue Reading »
Tags: apprendimento, cervello, farmaco, informazione, Mappa, Memoria, NMDAR, plasticità, spazio15 luglio 2010 - 3:35 pm
Il Coma (II Parte): Le parole che ingannano
Preambolo
Nel post precedente ci siamo occupati delle condizioni vitali che rendono discutibile il prelievo di organi da persone ancora in vita. In quei casi si può parlare di eutanasia o di omicidio a seconda del punto di vista. Oggi vedremo, invece, quando questo prelievo é possibile e soprattutto scopriremo che anche questo è un interessante argomento di neuroscienze.
Quando è possibile il prelievo di un organo?
 Senza tanti giri di parole, il prelievo é intuibilmente possibile quando il paziente é morto; tuttavia dopo il collasso cardio-respiratorio la maggior parte degli organi si deteriorano in maniera irreversibile e non possono essere riutilizzati. Si rende necessario, quindi, una particolare condizione di morte che conserva almeno parzialmente la vitalità degli organi; una condizione che é relativamente rara.
Senza tanti giri di parole, il prelievo é intuibilmente possibile quando il paziente é morto; tuttavia dopo il collasso cardio-respiratorio la maggior parte degli organi si deteriorano in maniera irreversibile e non possono essere riutilizzati. Si rende necessario, quindi, una particolare condizione di morte che conserva almeno parzialmente la vitalità degli organi; una condizione che é relativamente rara.
Per capire bene di cosa parliamo dobbiamo definire grossolanamente quando un paziente può essere definito ‘morto’.
Tutti sanno, almeno per via intuitiva, che un organo o una persona si possono considerare morti quando non è più possibile ripristinarne la normale attività e c’è un progressivo disfacimento delle strutture biologiche, ma in quale momento si muore effettivamente?
Secondo la più antica tradizione pagana e cattolica, un individuo muore quando cessa di battere il cuore (morte clinica), ma oggi sappiamo che uno shock elettrico può farlo ripartire, anche un trapianto o un sistema di pompe può ridare una vita relativamente normale ad una persona con un cuore morto. Secondo i mussulmani è il respiro ad essere la fonte della vita, la morte sopraggiungerebbe con l’ultima esalazione; ma anche in questo caso esistono delle persone in terapia intensiva che possono respirare grazie a delle macchine di ventilazione forzata.
In conclusione tutti questi casi comprendono solo delle formalità non sostanziali per identificare la morte. Possiamo, infatti, ripetere questa analisi organo per organo e non trovare un metodo certo che identifichi l’effettivo passaggio dalla vita alla morte. Lo stesso Karol Wojtyla, papa Giovanni Paolo II, aggiornò la definizione di morte per la chiesa con la separazione dell’anima dal corpo; un momento condivisibile ma difficilmente identificabile.
Negli anni, il nostro mondo burocratico ha classificato la morte secondo tre diversi criteri:
- morte clinica
- morte effettiva
- morte legale
La morte clinica
 La classificazione più nota é la cosiddetta morte clinica, ovvero uno stadio clinico precario dove il cuore non batte spontaneamente, a cui può seguire una vera morte se non si riesce ad intervenire efficacemente. In effetti si tratta solo di una definizione ingannevole il cui uso ed abuso dalla letteratura ha dato origine a miti e leggende di tutti i tipi per storie che colpiscono l’immaginario umano. Una persona ‘clinicamente morta’ è solo una persona che non manifesta dei chiari sintomi di vita come il battito cardiaco, la respirazione spontanea e la risposta a stimoli esterni. Tuttavia è noto che questo stadio non coincide con la vera morte poiché può essere indotto e revertito totalmente entro certi limiti temporali.
La classificazione più nota é la cosiddetta morte clinica, ovvero uno stadio clinico precario dove il cuore non batte spontaneamente, a cui può seguire una vera morte se non si riesce ad intervenire efficacemente. In effetti si tratta solo di una definizione ingannevole il cui uso ed abuso dalla letteratura ha dato origine a miti e leggende di tutti i tipi per storie che colpiscono l’immaginario umano. Una persona ‘clinicamente morta’ è solo una persona che non manifesta dei chiari sintomi di vita come il battito cardiaco, la respirazione spontanea e la risposta a stimoli esterni. Tuttavia è noto che questo stadio non coincide con la vera morte poiché può essere indotto e revertito totalmente entro certi limiti temporali.
Paragonare la morte clinica alla vera morte sarebbe come considerare morta una persona che non riesce a respirare. É ovvio che una persona in queste condizioni é ancora viva e, se si riesce a risolvere il problema in tempo, continua ad essere tale senza dover ‘resuscitare’. D’altra parte se il blocco respiratorio si prolunga troppo sopraggiunge un punto di non ritorno che possiamo considerare la morte effettiva.
La morte effettiva
La morte effettiva è il vero passaggio dalla vita alla morte ed è difficilmente identificabile con un unico istante. L’organismo, infatti, può morire tessuto per tessuto o organo per organo in tempi diversi, come anche il disfacimento delle strutture stesse. E’ noto che un paziente morto anche da diverse ore può donare le cornee, poiché le cellule da cui sono composte sopravvivono bene in assenza di ossigenazione, mentre diverso è il caso dei reni, poiché se non sono sufficientemente ossigenati e conservati, collassano entro pochi minuti.
L’argomento è ricco di riflessioni filosofiche, biologiche e pratiche, tuttavia rimane solo un concetto difficilmente identificabile.
La morte legale
 Se da un lato è difficile identificare quando avviene il passaggio dalla vita alla morte effettiva, perché gli organi del nostro corpo muoiono in tempi diversi, credo sia anche intuitivo che l’unico organo effettivamente importante per la nostra identità, la nostra memoria e la nostra coscienza sia il cervello. Morto un cuore, lo si può sostituire, così come anche i polmoni, i reni etc etc senza alterare la personalità dell’individuo; è evidente invece che quando muore il nostro cervello l’identità è irrimediabilmente persa. In queste condizioni, tutti i misteri, i ricordi e quello che può essere contenuto in ognuno di noi si disperde in una diffusa necrosi cerebrale, anche se il resto dell’organismo continua a vivere per un po’. Sottolineo che non si tratta solo di pura filosofia sull’identità della persona perché il cervello sovraintende anche una serie di funzioni fisiologiche come il controllo termico, pressorio, ormonale, metabolico, di crescita, di risposta immune etc etc. Senza la ‘sala comandi’ l’organismo per quanto curato in maniera intensiva morirà di lì a breve, poco a poco.
Se da un lato è difficile identificare quando avviene il passaggio dalla vita alla morte effettiva, perché gli organi del nostro corpo muoiono in tempi diversi, credo sia anche intuitivo che l’unico organo effettivamente importante per la nostra identità, la nostra memoria e la nostra coscienza sia il cervello. Morto un cuore, lo si può sostituire, così come anche i polmoni, i reni etc etc senza alterare la personalità dell’individuo; è evidente invece che quando muore il nostro cervello l’identità è irrimediabilmente persa. In queste condizioni, tutti i misteri, i ricordi e quello che può essere contenuto in ognuno di noi si disperde in una diffusa necrosi cerebrale, anche se il resto dell’organismo continua a vivere per un po’. Sottolineo che non si tratta solo di pura filosofia sull’identità della persona perché il cervello sovraintende anche una serie di funzioni fisiologiche come il controllo termico, pressorio, ormonale, metabolico, di crescita, di risposta immune etc etc. Senza la ‘sala comandi’ l’organismo per quanto curato in maniera intensiva morirà di lì a breve, poco a poco.
Per queste ragioni si definisce un individuo morto dal punto di vista legale, quando muore il suo cervello, anche se il suo corpo continua a vivere. A differenza della morte effettiva che è solo una definizione concettuale difficilmente identificabile, la morte legale, invece, è un concetto chiaro ed obiettivo. Le informazioni e le tecniche di indagine sulla morte cerebrale appartengono ad un importante campo neuroscientifico fatto di un intenso lavoro.
Le parole che ingannano
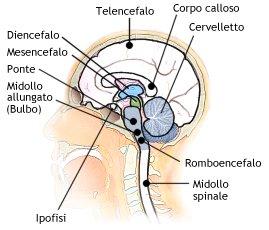
 Una volta chiarita per sommi capi la definizione di morte, può essere più chiara la definizione di coma irreversibile che si presenta con la morte cerebrale a cui segue la morte dell’organismo per le ragioni sopra descritte. In effetti il coma irreversibile è sommariamente sovrapponibile alla morte legale, le minime differenze possono essere tralasciate in questo post divulgativo.
Una volta chiarita per sommi capi la definizione di morte, può essere più chiara la definizione di coma irreversibile che si presenta con la morte cerebrale a cui segue la morte dell’organismo per le ragioni sopra descritte. In effetti il coma irreversibile è sommariamente sovrapponibile alla morte legale, le minime differenze possono essere tralasciate in questo post divulgativo.
Bisogna sottolineare che purtroppo il termine ‘coma irreversibile’ é alquanto ingannevole per i non addetti ai lavori, poiché contiene la parola coma che ricorda qualcuno ancora in vita, e la parola irreversibile che potrebbe indurre a pensare ad un coma molto lungo (vedi coma persistente).
A mettere benzina sul fuoco sono le centinaia, se non migliaia, di casi ‘miracolosi’ in cui il paziente é improvvisamente uscito da un lunghissimo coma di decine di anni. Le storie sono poi condite dalla mancanza di speranze dei medici seguita dallo stupore, preghiere, sogni premonitori ed ovviamente la mancanza di spiegazioni scientifiche. Quanta ignoranza in questa confusione. Nessuno sottolinea abbastanza, e ancor di meno, nessuno ricorda mai che tutti questi pazienti menzionati erano in coma persistente, già citato nel post precedente, ed il risveglio o l’evoluzione allo stato vegetativo sono il suo normale sviluppo.
Ovviamente il passaggio da un coma persistente ad un risveglio è un caso molto raro, per cui i medici non danno mai troppe illusioni ai parenti dei pazienti in questo stato. Quando questo succede c’è lo stupore di vedere un caso su un milione e la mancanza di spiegazione scientifica per il risveglio è data dalla mancanza di informazione su cosa tiene in coma una persona e cosa la può svegliare; nulla di miracoloso.
In definitiva mentre un coma persistente è una persona in stato di incoscienza a cui basterebbe un qualcosa, per ora ignoto, per tornare a rivivere una vita ‘normale’, il coma irreversibile é di fatto una tipologia morte. Basti pensare che per quanto intensamente curato, un coma irreversibile, non può mai arrivare ad anni di simil-vita, nella maggioranza dei casi l’organismo muore nel giro di ore, o giorni. Inutile rimarcare che nessun paziente si é mai svegliato da un coma irreversibile.
Il compito del medico è quello di identificare il coma e distinguerlo dal coma irreversibile (o morte legale) e comportarsi di conseguenza. Nel primo caso si assiste il paziente per quelle che sono le attuali conoscenze mediche, nel secondo caso si attiva la procedura della donazione degli organi.
Conclusione
In questo breve post abbiamo introdotto alcuni concetti di morte e li abbiamo distinti dal coma. Abbiamo anche visto che tutte le forme di coma, insieme alla morte clinica, non sono compatibili con la donazione degli organi poiché il donatore è ancora vivo. L’unica condizione che rende possibile la donazione degli organi è rappresentato dalla morte legale, o coma irreversibile, di un paziente che conserva ancora la vitalità degli organi. Questo è ovviamente un campo neuroscientifico di notevole interesse sotto molti aspetti (bioetico, medico, legale, scientifico, filosofico, etc)
Nel prossimo post, se vi farà piacere di leggerlo, vedremo a che punto è la ricerca neuroscientifica in questo campo e soprattutto cosa ci si aspetta dal futuro.
Alla prossima
Tags: coma, morte, Neuroscienze28 giugno 2010 - 7:28 pm
Il Coma (I Parte): Il Limbo della Coscienza
Preambolo
Un po’ di tempo fa mi é capitato di discutere con una persona dell’etica medica riguardo la donazione degli organi dei pazienti in coma. In particolare c’erano forti dubbi sul perché alcuni medici stacchino subito la spina per l’espianto degli organi, mentre in altri casi attendano anni. In seguito a questa discussione mi sono accorto ben presto che la maggioranza delle persone non ha una chiara idea di cosa sia effettivamente un coma e quando sia possibile espiantare gli organi. Non entro nel merito delle ragioni, ma credo che per avere una qualsiasi legittima opinione bisogna quantomeno essere certi di conoscere bene l’argomento. Tralasciando i pareri personali che sono propri ed inoppugnabili, ecco quanto segue in termini più o meno tecnici e divulgativi. A voi farvene una opinione.
Definizioni
 Come avrete sicuramente intuito, oggi parleremo di un argomento piuttosto difficile, portato alla più ampia diffusione da un evento di cronaca. La parola d’ordine di questo post sarà ‘coma‘, dal greco κῶμα (koma, sonno profondo), che si definisce semplicemente come un profondo stato di incoscienza simile al sonno che però non è suscettibile di risveglio. Da questa parola deriva poi lo stato comatoso che si definisce come un’apparente condizione simil-dormiente in cui l’individuo non è in grado di rispondere agli stimoli esterni.
Come avrete sicuramente intuito, oggi parleremo di un argomento piuttosto difficile, portato alla più ampia diffusione da un evento di cronaca. La parola d’ordine di questo post sarà ‘coma‘, dal greco κῶμα (koma, sonno profondo), che si definisce semplicemente come un profondo stato di incoscienza simile al sonno che però non è suscettibile di risveglio. Da questa parola deriva poi lo stato comatoso che si definisce come un’apparente condizione simil-dormiente in cui l’individuo non è in grado di rispondere agli stimoli esterni.
Le definizioni sono alquanto generiche perché comprendono vari stadi di gravità (profondità del coma) e dobbiamo definire ulteriori limiti per capire meglio.
Cosa NON è il Coma
Ci sono 5 casi che rispondono apparentemente allo stato comatoso ma non sono tali:
- Il sonno non è coma poiché lo stato di incoscienza è solo parziale e facilmente reversibile con un rumore, con il tatto o con un’abitudine.
- La morte che appare come la suddetta definizione, ma é una situazione evidentemente diversa.
- La sindrome locked-in, in cui i muscoli volontari dell’individuo sono completamente paralizzati, e non c’é possibilità di interagire con l’ambiente. Il paziente è pienamente cosciente di sé, sveglio e vigile, ma letteralmente immobile.
- Lo stato vegetativo, dove il paziente appare sveglio, ma non risponde agli stimoli esterni. Le funzioni di base come la respirazione, il ciclo biologico di sonno veglia restano intatte. Raramente possono anche afferrare degli oggetti in maniera istintiva.
- Lo stato stuporoso, un effetto transiente simile allo stato vegetativo, dovuto generalmente ad uno shock. Può essere facilmente interrotto o revertito mediante stimoli che attivino meccanismi istintivi ed irrazionali (es. pizzicotto).
Cosa è il Coma
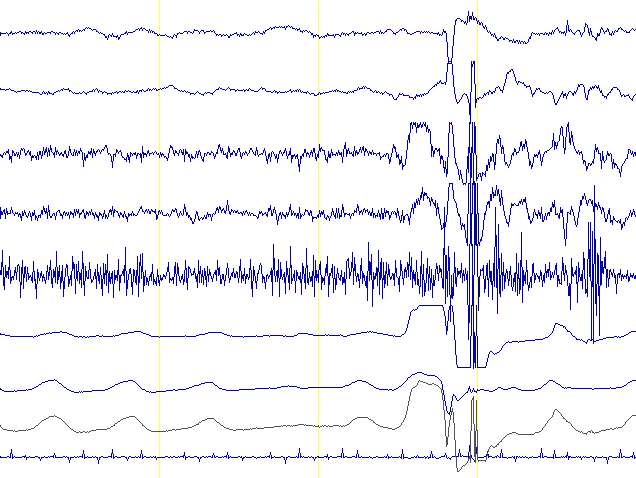
Una volta definito cosa non è il coma, ora passiamo a capire meglio quale condizione possa essere definita tale.
Nel coma abbiamo una scissione della coscienza dal mondo esterno in maniera simile ad un’anestesia generale. Il meccanismo esatto che spegne i canali di comunicazione tra alcune zone del cervello, e che causa lo stato comatoso, è tutt’ora ignoto; si sa solo che è implicato il danneggiamento o l’inibizione della regione reticolare del romboencefalo. Questa zona del cervello, tra le altre cose, partecipa al ciclo sonno-veglia ed indirettamente partecipa allo ‘spegnimento’ della coscienza durante il sonno. Forse é per questo meccanismo condiviso che nel coma il cervello può percepire incoscientemente il mondo esterno in maniera simile a quanto avviene durante il sonno. Tuttavia, la differenza più significativa tra un paziente che dorme ed uno in coma è che quest’ultimo potrebbe subire un’operazione chirurgica senza rendersene conto e soprattutto senza svegliarsi.
C’é coma e coma
 Non tutti i coma sono identici così come non sono simili le cause scatenanti. La metodica più elementare per valutare la ‘profondità’ del coma è la Scala di Glasgow (GCS) che si basa sulla risposta agli stimoli oculari, verbali e motori. Ad ognuno di questi viene assegnato un punteggio la cui somma costituisce l’indice GCS. L’indice può andare da 3 (coma profondo) a 15 (paziente sveglio e cosciente).
Non tutti i coma sono identici così come non sono simili le cause scatenanti. La metodica più elementare per valutare la ‘profondità’ del coma è la Scala di Glasgow (GCS) che si basa sulla risposta agli stimoli oculari, verbali e motori. Ad ognuno di questi viene assegnato un punteggio la cui somma costituisce l’indice GCS. L’indice può andare da 3 (coma profondo) a 15 (paziente sveglio e cosciente).
Un paziente in coma può ripetutamente svegliarsi, guardare intorno, afferrare qualcosa, magari farfugliare qualche parola e persino muoversi, senza però potersi definire fuori dal coma. C’é da dire, però, che tutti questi sintomi non sono innescati da eventi esterni come ad esempio un rumore, la vista di un oggetto etc, e neppure da un apparente ciclo spontaneo. Ogni condizione fisiologica guidata dall’encefalo é soggetto a stimoli apparentemente casuali ed imprevedibili, persino la respirazione in queste persone è soggetto a discontinuità. E’ da sottolineare che l’individuo è vivo, ha una coscienza, la sua memoria e tutto quanto sia parte della sua personalità resta intatta; c’è solo un problema nel ‘svegliarlo’ o comunque collegare la sua coscienza con gli stimoli esterni.
Cosa succede ‘dopo’?
 Una volta inquadrata la patologia per schemi intuitivi ed in maniera molto generale ora vediamo cosa può portare al coma e cosa avviene dopo.
Una volta inquadrata la patologia per schemi intuitivi ed in maniera molto generale ora vediamo cosa può portare al coma e cosa avviene dopo.
Tra le cause più comuni che possono portare al coma ci sono le intossicazioni (stupefacenti, alcool, tossine etc), le alterazioni del metabolismo (ipoglicemia, iperglicemia, chetoacidosi) o danni e malattie del sistema nervoso centrale (ictus, traumi cranici, ipossia, edema etc). L’induzione in coma, specie se traumatico, può portare direttamente alla morte nella fase acuta, oppure può seguire due vie più o meno lunghe:
- recupero della coscienza con un ritorno parziale o totale alla normale vita quotidiana
- evolvere nello stato vegetativo.
Tranne per il coma post-traumatico e quello indotto farmacologicamente, lo stato comatoso raramente persiste per più di 4 settimane, e per lo più il ripristino delle normali facoltà mentali è spontaneo. Se ne deduce che il risveglio è molto probabile nelle prime 4 settimane, anche se il recupero può non essere totale. Quando il coma supera abbondantemente le 4 settimane si definisce ‘coma persistente’ poiché è noto che le probabilità di risveglio spontaneo diventano molto scarse, anche se non impossibili. La maggioranza delle persone ‘miracolosamente’ uscite dal coma, infatti, appartengono a questo gruppo.
Ad ogni modo dobbiamo sottolineare che un paziente in coma NON è morto e quindi non è possibile effettuare alcun espianto di organi. Il risveglio, seppur estremamente improbabile, è ancora possibile come anche il ritorno ad una vita ‘normale’.
Lo stato vegetativo
 Purtroppo l’alternativa più frequente al ritorno ad una vita normale dal coma è l’evoluzione allo stato vegetativo. Questa complessa situazione, già definita prima in maniera intuitiva, é facilmente distinguibile dal coma stesso. Nello stato vegetativo, infatti, le funzioni biologiche tornano ad una ‘apparentemente normalità’, l’individuo mostra uno spontaneo ciclo di sonno veglia, può osservare degli oggetti intorno, ed in alcuni casi può camminare, piangere, ridere ed altro. Raramente ci possono essere delle condizioni esterne che possono alterare queste funzioni biologiche.
Purtroppo l’alternativa più frequente al ritorno ad una vita normale dal coma è l’evoluzione allo stato vegetativo. Questa complessa situazione, già definita prima in maniera intuitiva, é facilmente distinguibile dal coma stesso. Nello stato vegetativo, infatti, le funzioni biologiche tornano ad una ‘apparentemente normalità’, l’individuo mostra uno spontaneo ciclo di sonno veglia, può osservare degli oggetti intorno, ed in alcuni casi può camminare, piangere, ridere ed altro. Raramente ci possono essere delle condizioni esterne che possono alterare queste funzioni biologiche.
Alcuni definiscono questi pazienti come dei gusci vuoti senza volontà e senza sentimenti. In alcuni casi questo corrisponde ad una obiettiva realtà, come ad esempio nelle persone che hanno subito un grave danneggiamento cerebrale delle funzioni superiori. Tuttavia ciò non é vero per tutti gli altri pazienti.
Anche in questo caso il ritorno miracoloso ad una vita ‘normale’ dopo lo stato vegetativo è relativamente raro ma non impossibile. Chi è affascinato dalla ricerca e dalla filmografia potrebbe vedere il film “Risvegli” con Robin Williams e Robert De Niro, basato su fatti realmente accaduti, che esplicano bene il risveglio di un gruppo di persone da uno stato vegetativo a cui erano state tolte tutte le speranze da tantissimi anni. Purtroppo ancora oggi non abbiamo ancora trovato un metodo efficace ed universale per sbloccare il corto circuito cerebrale che avviene in questi pazienti, ma c’é sempre la speranza di ritrovare un metodo in futuro.
C’é da dire che l’espianto di organi equivale ad uccidere il paziente o ad una eutanasia, secondo il vostro punto di vista, per cui il medico é impossibilitato a ‘staccare la spina’ per espiantare gli organi.
Conclusioni
In questo breve post introduttivo abbiamo definito il coma per via intuitiva ed esaminato alcune condizioni tratte dall’esperienza senza entrare nel merito dell’etica. Abbiamo visto anche che l’espianto di organi di questi pazienti è alquanto discutibile poiché non sono ancora morti e in opportune condizioni potrebbero essere recuperati.
Nel prossimo post, invece, vedremo quali sono le condizioni che la medicina considera opportune per l’espianto degli organi.
Tags: coma, Ictus, Percezione, stato comatoso, stato vegetativo16 giugno 2010 - 5:25 pm
Le cellule-stella sono tante, milioni di milioni…Il responso finale (O quasi)
 A cosa servono gli astrociti? Negli scorsi post abbiamo cercato di rispondere a questo interrogativo esaminando due opinioni opposte. Chi dice che le cellule-stella abbiano un ruolo nei processi cognitivi superiori come apprendimento e memoria, e chi dice che queste cellule siano solo di supporto ai neuroni. Qual’è la verità? Ad oggi, non abbiamo ancora certezze. Però, esiste un possibile “modello” che spiega quanto si è finora scoperto. Continue Reading »
A cosa servono gli astrociti? Negli scorsi post abbiamo cercato di rispondere a questo interrogativo esaminando due opinioni opposte. Chi dice che le cellule-stella abbiano un ruolo nei processi cognitivi superiori come apprendimento e memoria, e chi dice che queste cellule siano solo di supporto ai neuroni. Qual’è la verità? Ad oggi, non abbiamo ancora certezze. Però, esiste un possibile “modello” che spiega quanto si è finora scoperto. Continue Reading »
31 maggio 2010 - 7:32 pm
Memoria e astrociti: una buia notte senza stelle?
 Memoria, apprendimento… Meccanismi ben lontani dall’essere svelati. I neuroni, fino a pochi anni fa dominatori indiscussi della scena, stanno dividendo le luci della ribalta con qualcuno che potrebbe togliere loro il ruolo di protagonisti nei fenomeni neurologici: gli astrociti. Le ormai famose cellule-stella sono al centro di una diatriba: c’è chi sostiene un loro ruolo nella comunicazione tra neuroni e chi al contrario vorrebbe continuare a considerare queste cellule come “assistenti” che aiutano le cellule neuronali a vivere ben nutrite.
Memoria, apprendimento… Meccanismi ben lontani dall’essere svelati. I neuroni, fino a pochi anni fa dominatori indiscussi della scena, stanno dividendo le luci della ribalta con qualcuno che potrebbe togliere loro il ruolo di protagonisti nei fenomeni neurologici: gli astrociti. Le ormai famose cellule-stella sono al centro di una diatriba: c’è chi sostiene un loro ruolo nella comunicazione tra neuroni e chi al contrario vorrebbe continuare a considerare queste cellule come “assistenti” che aiutano le cellule neuronali a vivere ben nutrite.
Quale sarà la giusta soluzione? Continue Reading »
Tags: Astrociti, Calcio, Cellule-stella, GPCR, IP3, Memoria, Neuroni, plasticità neuronale, recettori, sinapsi31 maggio 2010 - 10:29 am
Quando il Cervello Decide il Suicidio (VIII Parte): Gli 007 del SNC
Gli 007 del Sistema Nervoso Centrale
Preambolo
Eccoci di nuovo con il seguito del nostro giallo misterioso, ed iniziamo subito con un breve riassunto.
Siamo nei panni di un improbabile investigatore molecolare che deve scoprire chi ha causato la morte dei neuroni in seguito ad ictus. Nelle prime puntate abbiamo appreso la complessità dell’evento ischemico, ciò che ne consegue, e la volontà preprogrammata di qualcuno di assassinare i neuroni. Nella puntata precedente, invece, abbiamo introdotto le cellule microgliali che rappresentano delle guardie giurate dedicate alla protezione del cervello. Oggi vedremo come queste cellule siano più simili a degli agenti segreti.
L’interrogatorio
Per istituire un processo contro qualcuno é necessario avere delle prove convincenti; i sospetti da soli non bastano.
Ecco i fatti:
- Il cervello si trova in una condizione isolata, prima, durante e dopo il neuroomicidio;
- C’è sicuramente un colpevole e deve necessariamente essere contenuto all’interno del cervello;
- In questa ‘stanza’ troviamo permanentemente i neuroni (le principali vittime), la glia (vittime collaterali), e la microglia (guardie giurate);
- La microglia, il terzo incomodo, sopravvive al danno e si allontana dai neuroni poco prima che muoiano;
- La microglia in opportune condizioni riesce ad uccidere le cellule patogene ed anche i neuroni in agonia;
- La microglia non ha ancora un alibi per l’evento ischemico.
Continuiamo quindi l’interrogatorio nei confronti della microglia.

“…in altre parole i neuroni sono morti poco dopo la sua visita?”
“Sì, ma ero già lontano quando questo è successo”
“Può spiegarci perché Lei si è allontanato dal danno neuronale salvandosi la pelle, invece che combattere fino all’ultimo respiro? Dopotutto questo è il Suo ruolo, Lei è una guardia giurata e non può allontanarsi quando le cose vanno male”
“In realtà c’era ben poco da fare quando sono arrivato sul luogo del delitto, i neuroni stavano già morendo, non ho potuto far nulla ed era inutile sacrificarsi. Noi cellule microgliali non abbiamo alcuna possibilità di rinforzo da parte di progenitori esterni alla BEE, quindi sacrificarsi durante l’ischemia avrebbe comportato una perdita di cellule con una conseguente perdita di protezione per gli altri neuroni ancora vivi.”
“…uhmm… se ho capito bene, Lei è arrivato in ritardo, non ha fatto nulla e si è allontanato quando le cose si sono messe male. Perchè?”
“Sono preparato per affrontare cellule patogene, infezioni, infiammazioni ed eventi simili. L’evoluzione non mi ha selezionato per affrontare l’assenza di ossigeno e nutrienti.”
“…e cosa ha fatto in quei brevi momenti in cui Lei era a contatto con tanti neuroni poco prima che morissero?”
“Ho liberato dei fattori trofici per sostenere i neuroni e la glia. Generalmente i neuroni sono meno sensibili alla morte quando ricevono degli ormoni. Il mio ruolo era quello di incoraggiarli a vivere fino a quando fosse possibile”
Sembra non esserci nulla di serio, ma il dubbio rimane; ad esempio come mai una cellula tanto efficiente nel riconoscere i danni neuronali si è accorta del danno in corso così tanto in ritardo e non abbia potuto far nulla? La morte neuronale sembra avvenire poco dopo la scomparsa della microglia, può essere tutto questo un caso?
La ricostruzione del delitto
 Prima di trarre conclusioni affrettate andiamo in laboratorio con la nostra ricostruzione della scena del crimine. Induciamo una ischemia cerebrale a dei ratti ed analizziamo nel tempo cosa fa la microglia nel SNC. Anche se non è facile seguire le cellule microgliali che girano nel cervello quando l’animale è vivo, è possibile ricostruire la scena mettendo insieme una serie di animali analizzati a tempi diversi dopo l’evento ischemico.
Prima di trarre conclusioni affrettate andiamo in laboratorio con la nostra ricostruzione della scena del crimine. Induciamo una ischemia cerebrale a dei ratti ed analizziamo nel tempo cosa fa la microglia nel SNC. Anche se non è facile seguire le cellule microgliali che girano nel cervello quando l’animale è vivo, è possibile ricostruire la scena mettendo insieme una serie di animali analizzati a tempi diversi dopo l’evento ischemico.
Qui la prima sorpresa. Nel SNC tutte le cellule neuronali e astrocitarie sono pressoché fisse, magari con una mobilità molto limitata e lenta, mentre le cellule microgliali si muovono ad una velocità impressionante. Quello che colpisce molto è la capacità di attraversare reti di assoni, di dendrociti, cellule neuronali ed astrocitarie senza causare danno e senza avere punti di appiglio.
La seconda sorpresa è ancora più scioccante, la forma, le dimensioni e le caratteristiche di queste cellule è enormemente vasta, difficile riconoscerle anche per un occhio esperto. Si possono definire almeno 7 fenotipi principali di microglia: ameboide, ramificata, attiva e non fagocitica, attiva e fagocitica, cellule del composto granulare, perivascolare e juxatavascolare.
Neanche uno 007 potrebbe possedere altrettante identità. Per seguirle, quindi, possiamo usare dei markers, generici come Iba-1 o RCA-1 per riconoscere un po’ tutte le identità microgliali, oppure selettivi come ED1 per riconoscere solo le cellule microgliali attivate. Vi renderete subito conto della complessità di tale analisi, poiché nessun marker fino ad ora scoperto è effettivamente in grado di riconoscere tutte le identità microgliali in maniera specifica e selettiva. Insomma queste cellule sono qualcosa di sfuggente come potrebbe essere un agente segreto.
Ad aggiungere ulteriori sospetti c’è il fatto che le cellule microgliali si avvicinano ai neuroni con una certa identità e se ne allontanano con un’altra. Se poi pensiamo di fare una perquisizione e cercare cosa trasportano le cellule microgliali nelle vescicole potremmo ritrovare una serie infinita di endosomi, lisosomi, autosomi, e vescicole che contengono sostanze altamente tossiche, materiali infiammabili insieme a sostanze neurotrofiche etc etc. Di certo non è quello che si ritroverebbe in una borsa di un medico o infermiere.
Il Processo: L’accusa
 L’accusa inizia l’arringa con delle frasi taglienti e tendenziose. Secondo la sua ricostruzione le cellule microgliali, avvezze all’eutanasia, si sono dirette verso la zona sofferente dall’ipossia. I dati pubblici ci hanno depistato sull’effettivo orario di arrivo sul luogo del delitto, in realtà, recenti scoperte ci indicano che le cellule microgliali si trovavano già in loco prima del tempo dichiarato. Non è stato possibile vederle poiché sono arrivate con un travestimento non noto fino a poco tempo fa. Poi una volta arrivate sul luogo prefissato hanno iniziato a contornare alcuni neuroni in sofferenza. La microglia, adese ai neuroni ha liberato delle sostanze in uno spazio estremamente piccolo, troppo scarse per essere analizzate e troppo rapide per essere viste. In questo istante forse c’é stato un ‘errore’, ed al posto di sostanze neurotrofiche la microglia ha lasciato delle sostanze tossiche. Magari ha valutato che i neuroni non ce l’avrebbero fatta a passare indenni l’ipossia e li ha avvelenati in maniera irreversibile per eutanasia. Poi mentre i neuroni avvelenati stavano resistendo all’ipossia, il colpo di grazia, la microglia si é allontanata aggravando la situazione per il mancato sostentamento trofico. Ciò che ne é conseguito é la morte dei neuroni in circostanze apparentemente solitarie. Poi sono arrivate le cellule microgliali travestite da spazzine ed hanno pulito il luogo del delitto.
L’accusa inizia l’arringa con delle frasi taglienti e tendenziose. Secondo la sua ricostruzione le cellule microgliali, avvezze all’eutanasia, si sono dirette verso la zona sofferente dall’ipossia. I dati pubblici ci hanno depistato sull’effettivo orario di arrivo sul luogo del delitto, in realtà, recenti scoperte ci indicano che le cellule microgliali si trovavano già in loco prima del tempo dichiarato. Non è stato possibile vederle poiché sono arrivate con un travestimento non noto fino a poco tempo fa. Poi una volta arrivate sul luogo prefissato hanno iniziato a contornare alcuni neuroni in sofferenza. La microglia, adese ai neuroni ha liberato delle sostanze in uno spazio estremamente piccolo, troppo scarse per essere analizzate e troppo rapide per essere viste. In questo istante forse c’é stato un ‘errore’, ed al posto di sostanze neurotrofiche la microglia ha lasciato delle sostanze tossiche. Magari ha valutato che i neuroni non ce l’avrebbero fatta a passare indenni l’ipossia e li ha avvelenati in maniera irreversibile per eutanasia. Poi mentre i neuroni avvelenati stavano resistendo all’ipossia, il colpo di grazia, la microglia si é allontanata aggravando la situazione per il mancato sostentamento trofico. Ciò che ne é conseguito é la morte dei neuroni in circostanze apparentemente solitarie. Poi sono arrivate le cellule microgliali travestite da spazzine ed hanno pulito il luogo del delitto.
L’accusa continua rimarcando più volte il passato della microglia, come una cellula già avvezza al neuroomicidio (precedenti penali), inoltre ha mentito sull’orario di arrivo (depistaggi), si trovava vicino ai neuroni durante i fatti (opportunità), aveva dentro di sé l’arma del delitto (capacità), poteva girovagare indisturbato per il cervello con una discreta velocità (possibilità), si è allontanata con un travestimento prima che i neuroni si riprendessero causandone la morte (mancato soccorso). Ed in fine, l’arringa termina con una frase lapidaria: come nei più classici e proverbiali gialli, l’assassino è ritornato sul luogo del delitto ed ha rimosso le prove per fagocitosi (occultamento delle prove).
Non si tratta di un evento accidentale, ma di un pluriomicidio aggravato e premeditato di neuroni ed astrociti da parte di ‘servizi segreti deviati’ appartenenti al SNC.
La parola alla difesa
 La difesa risponde punto per punto con calma e freddezza.
La difesa risponde punto per punto con calma e freddezza.
Le multi identità sono parte del lavoro fisiologico della microglia. Queste cellule, infatti, in mancanza di collaboratrici, (es macrofagi, cellule presentanti l’antigene, cellule B etc,) devono necessariamente fare tutto da sole adattandosi di volta in volta alle nuove necessità. Sull’orario non c’é stata alcuna dichiarazione falsa, ogni cellula della microglia esprime o meno degli antigeni in funzione del lavoro che sta facendo, se non esistono ancora metodi per marcarli tutte ciò non indica alcuna colpevolezza.
Sulla questione delle sostanze tossiche e letali, é ovvio che una guardia giurata abbia sempre con sé la pistola ed il caricatore sempre pronti, tuttavia può anche avere con sé i materiali di primo soccorso. Fino ad ora non ci sono prove che la microglia abbia liberato sostanze tossiche verso i neuroni in quei momenti.
Anzi, l’imputato si è dimostrato utile per rimuovere le sostanze potenzialmente tossiche, ovvero quei residui post-ischemici che aumentano l’infiammazione e la sofferenza neuronale. Inoltre la microglia ha creato anche un ambiente idoneo per la formazione di nuove sinapsi per la ripresa post-danno.
La difesa poi ricorda che i topi transgenici che non hanno la microglia non sopravvivono all’embriogenesi, per cui stiamo trattando di cellule essenziali per la vita stessa. Poi cita gli innumerevoli casi in cui la microglia ha svolto ruoli eroici in difesa dei nostri delicati neuroni. Tutto questo non può essere cancellato da qualche sospetto e soprattutto da condizioni alle quali la microglia stessa non era preparata.
Se questo fosse un giallo…
 Se questo fosse un giallo non potrebbe mancare il classico colpo di scena. Questa volta arriva proprio da Mario Capecchi che scopre, in una sua recente pubblicazione su cell, che l’inattivazione della microglia provoca un fenotipo ossessivo-convulsivo. Quello che sorprende é che l’iniezione nel midollo osseo di cellule microgliali marcate e fluorescenti provoca la comparsa di cellule microgliali nel cervello. Ancora una volta la microglia ha mentito, e a quanto pare ha un passaggio segreto per oltrepassare la BEE, anche se oggi non é stato ancora possibile scoprire come. A quanto pare la microglia ha molti aspetti ancora oscuri, forse anche delle identità e dei passaporti che non conosciamo ancora.
Se questo fosse un giallo non potrebbe mancare il classico colpo di scena. Questa volta arriva proprio da Mario Capecchi che scopre, in una sua recente pubblicazione su cell, che l’inattivazione della microglia provoca un fenotipo ossessivo-convulsivo. Quello che sorprende é che l’iniezione nel midollo osseo di cellule microgliali marcate e fluorescenti provoca la comparsa di cellule microgliali nel cervello. Ancora una volta la microglia ha mentito, e a quanto pare ha un passaggio segreto per oltrepassare la BEE, anche se oggi non é stato ancora possibile scoprire come. A quanto pare la microglia ha molti aspetti ancora oscuri, forse anche delle identità e dei passaporti che non conosciamo ancora.
Inoltre come può l’iniezione midollare di una microglia inattiva causare un fenotipo psichiatrico? Siamo di fronte ad un personaggio davvero interessante.
Il giudizio
 Mettere insieme tutti questi dati sparsi non é semplice. Nella ricerca, infatti, é difficile soprattutto distinguere la verità obiettiva dalle interpretazioni, ed in questo lungo processo sono state fatte solo delle allusioni, niente di più. L’articolo di Capecchi parla di una interferenza sui recettori nocicettivi periferici da parte della microglia. Un topo con difficoltà nella percezione del dolore ha dei comportamenti anomali, ma non c’é niente di significativamente neurologico.
Mettere insieme tutti questi dati sparsi non é semplice. Nella ricerca, infatti, é difficile soprattutto distinguere la verità obiettiva dalle interpretazioni, ed in questo lungo processo sono state fatte solo delle allusioni, niente di più. L’articolo di Capecchi parla di una interferenza sui recettori nocicettivi periferici da parte della microglia. Un topo con difficoltà nella percezione del dolore ha dei comportamenti anomali, ma non c’é niente di significativamente neurologico.
La corte, quindi propone una nuova perizia. Verifichiamo cosa avviene in un topo transgenico capace di inattivare la funzione microgliale in maniera condizionata.
Il risultato é sconcertante, l’inattivazione condizionale della microglia, poco prima e durante l’ictus, provoca un notevole peggioramento del danno ischemico ed una sopravvivenza neuronale ridotta. Ovviamente ci sono state anche le critiche su quale gene é stato scelto per disattivare la microglia. Forse é stato scelto quel gene che sovraintende le funzioni microgliali ‘buone’. Si vedrà.
Tratta la somma, l’imputato é assolto per insufficienza di prove.
Conclusioni
Purtroppo, siamo ancora lontani dalla verità. Le indagini sulla microglia sono tutt’altro che concluse, si tratta di un campo in rapida espansione ed é sotto indagine per tantissime patologie come l’HIV, la sclerosi multipla, l’Alzheimer e tante altre patologie neurodegenerative.
Per ora il processo sull’ischemia é fermo per insufficienza di prove, ma qualcuno nel mondo sta ancora tentando di incastrare la microglia, chissà se ci sarà nuovamente qualche altro colpo di scena che la riporterà nuovamente sotto processo.
La ricerca, come al solito, non ha mai fine.
Alla prossima
Tags: Animali transgenici, Glia, Ictus, Ischemia Cerebrale, microglia, NeuroniPubblicità
- Sei interessato a promuovere la tua attività su Inside Research?
Pubblicità su MolecularLab.it.
Ultimi commenti
- L’Orecchio: Assoluto o Relativo? (Parte II) (2)
- di gaetano alle 11:21 am
- Di che colore è il numero 6? (40)
- di roxana alle 12:18 am
- Oligodendrociti e mielina. Se non è solo questione di materia grigia… (7)
- di giuseppe di virgilio alle 3:00 pm
- Incontro con la memoria (7)
- di terfle alle 12:11 pm
- Essere Anormali: Vantaggio o Svantaggio? (24)
- di Neuroscience alle 2:27 pm
- L’Orecchio: Assoluto o Relativo? (Parte II) (2)
Archivi
- giugno 2011
- aprile 2011
- marzo 2011
- novembre 2010
- ottobre 2010
- luglio 2010
- giugno 2010
- maggio 2010
- aprile 2010
- gennaio 2010
- dicembre 2009
- settembre 2009
- maggio 2009
- febbraio 2009
- gennaio 2009
- dicembre 2008
- novembre 2008
- settembre 2008
- agosto 2008
- luglio 2008
- maggio 2008
- aprile 2008
- marzo 2008
- febbraio 2008
- gennaio 2008
- dicembre 2007
- novembre 2007
Meta