4 settembre 2009 - 1:00 pm
Preambolo
Finalmente le tanto meritate vacanze sono iniziate anche per me, e colgo l’occasione per finire finalmente questo post in elaborazione oramai da tanto tempo, come al solito cortesemente corretto da Valentina. Qui descriverò un argomento abbastanza lungo, difficile e ‘spigoloso’ per i profani ed esperti, anche se penso che tutti lo conoscono almeno un po’ per sentito dire.
Parleremo dell’olfatto, un argomento non facile da raccontare in un post divulgativo senza ‘smussare’ qua e là il mosaico di teorie e di esperimenti che sono stati fatti fino ad ora.
L’Olfatto: Alcuni dettagli ovvi

Come al solito mi servirò inizialmente di qualche punto di riferimento per metterci d’accordo ed inizierò dagli odori che sono causati, come tutti voi sapete, da molecole sprigionate dalle sostanze e disperse nell’aria che respiriamo, dove raggiungono il nostro naso. L’altro punto di riferimento utile si trova subito dietro il naso e sopra la volta della nostra cavità buccale dove passano tutte le molecole che noi respiriamo. E’ una cavità rivestita da una mucosa di circa 10 cm2 che attraverso la solubilizzazione, la dispersione, l’adsorbimento e quant altro, ferma una porzione delle molecole che si trovano nello stadio aeriforme nella cavità nasale e le disperde nel fluido mucoso. Qui le molecole possono incontrare una o più cellule recettoriali olfattive (ORC), in un uomo adulto se ne contano circa 10-20 milioni, ricche di propagini dette ciglia e di proteine dette recettori olfattivi (da qui chiamate Odorant Receptor OR).
I OR hanno il compito ‘legare’ in maniera più o meno specifica alcuni gruppi funzionali di queste molecole presenti nel fluido mucoso, ed innescare una cascata di eventi che si amplifica fino ad attivare la cellula recettrice e scatenare la propagazione del ‘segnale odoroso’. Il cervello, a sua volta, provvederà ad interpretare il segnale in diversi stadi con una serie di meccanismi molto complessi. Per dettagli ed approfondimenti potete guardare questo link.
Fin qui il tutto è quasi scontato ed ovvio, però dalla teoria alla pratica c’è una notevole differenza, vi dico subito che ancora oggi ci sono vari lati oscuri che cercherò di descrivervi in parole semplici, portandovi ai problemi scientifici in maniera intuitiva per quanto sia nelle mie capacità.
Quanti odori esistono?
 Come penso molti di voi possono intuire, esiste un numero inimmaginabile di odori, e la maggioranza di questi è causato da un mix di ‘odori primari’ in diversi rapporti, ma a complicare la situazione c’è che una molecola odorosa può agire su decine di recettori diversi con diversa specificità. I OR dei composti aminici, ad esempio, riconoscono solamente l’azoto primario, secondario o terziario con diversa efficacia, per cui gli odori di questi composti non possono essere discriminati con facilità e sembrano tutti molto simili. C’è da dire però che nonostante questa piccola sovrapposizione di segnali si possono discriminare con elevata sensibilità circa 10.000 odori diversi.
Come penso molti di voi possono intuire, esiste un numero inimmaginabile di odori, e la maggioranza di questi è causato da un mix di ‘odori primari’ in diversi rapporti, ma a complicare la situazione c’è che una molecola odorosa può agire su decine di recettori diversi con diversa specificità. I OR dei composti aminici, ad esempio, riconoscono solamente l’azoto primario, secondario o terziario con diversa efficacia, per cui gli odori di questi composti non possono essere discriminati con facilità e sembrano tutti molto simili. C’è da dire però che nonostante questa piccola sovrapposizione di segnali si possono discriminare con elevata sensibilità circa 10.000 odori diversi.
Ciò premesso, consentitemi da questo punto in poi di semplificare e di concentrarci esclusivamente sugli ‘odori primari‘ che sono riconosciuti quasi esclusivamente da un solo tipo di OR e che danno una sensazione odorosa molto precisa.
Cosa c’è di difficile?
 Nel mondo animale, il non saper distinguere l’odore di una preda dall’odore del proprio predatore fa la differenza tra fare una buona colazione ed essere la colazione di qualcun altro. Anche riconoscere l’odore delle cose che si possono mangiare da quelle che è meglio evitare è fondamentale per la sopravvivenza, quindi i mammiferi durante l’evoluzione hanno accumulato più di 1.000 geni che sovraintendono il più misterioso dei 5 sensi. In pratica più del 3% del genoma dei mammiferi è deputato alla percezione degli odori, e dato che questi recettori possono riconoscere anche più tipologie di molecole con diversa affinità determinando una combinazione di interazioni intermedie, questo amplifica esponenzialmente il ventaglio di sensazioni che possono dare le molecole presenti nell’aria.
Nel mondo animale, il non saper distinguere l’odore di una preda dall’odore del proprio predatore fa la differenza tra fare una buona colazione ed essere la colazione di qualcun altro. Anche riconoscere l’odore delle cose che si possono mangiare da quelle che è meglio evitare è fondamentale per la sopravvivenza, quindi i mammiferi durante l’evoluzione hanno accumulato più di 1.000 geni che sovraintendono il più misterioso dei 5 sensi. In pratica più del 3% del genoma dei mammiferi è deputato alla percezione degli odori, e dato che questi recettori possono riconoscere anche più tipologie di molecole con diversa affinità determinando una combinazione di interazioni intermedie, questo amplifica esponenzialmente il ventaglio di sensazioni che possono dare le molecole presenti nell’aria.
La chiave di volta per ottenere un buon olfatto è sicuramente il collegamento tra le cellule recettoriali odorose (ORC) ed il cervello, ovvero quel meccanismo che consente di poter capire quale specifico OR è stato attivato. Si tratta di un meccanismo per nulla banale, e come punto di riferimento, possiamo considerare l’esempio del gusto, dove i recettori cellulari sono generati in base alla localizzazione sulla superficie della lingua. Nel cervello si forma una sorta di correlazione tra aree specifiche corticali ed aree della lingua denominata ‘mappa sensoriale‘. A questo consegue che è possibile percepire il salato, l’amaro, il dolce ed l’acido in base a quale zona della lingua è stata stimolata maggiormente.
Possiamo trasportare questa ipotesi nell’olfatto, ed immaginare che l’area in cui si trovano le ORC determina anche il set di recettori da esprimere. Ipotizziamo che ciascuna cellula recettrice dell’olfatto possa esprimere un centinaio o più OR localizzati in diverse aree della mucosa nasale. Ogni area in cui vi sono le ORC potrebbe essere correlata ad una zona del cervello a cui corrisponderebbero determinate sensazioni odorose.
Quanti recettori per cellula?
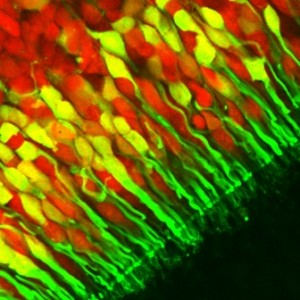 Sapere quanti tipi di recettori sono presenti per cellula è fondamentale per capire quante aree ci dovremmo aspettare di trovare e per verificarlo potremmo ricorrere a tecniche avanzatissime di in situ hybridization, e di retrotrascrizione ed amplificazione per PCR su singola cellula. Ovviamente non sono tecniche facili e nemmeno tanto diffuse, ma gli esperti sono riusciti a dimostrare con questi esperimenti che ogni ORC esprime solamente un singolo OR.
Sapere quanti tipi di recettori sono presenti per cellula è fondamentale per capire quante aree ci dovremmo aspettare di trovare e per verificarlo potremmo ricorrere a tecniche avanzatissime di in situ hybridization, e di retrotrascrizione ed amplificazione per PCR su singola cellula. Ovviamente non sono tecniche facili e nemmeno tanto diffuse, ma gli esperti sono riusciti a dimostrare con questi esperimenti che ogni ORC esprime solamente un singolo OR.
Una cosa simile avviene durante il differenziamento cellulare, dove la combinazione di alcuni fattori trascrizionali decide l’attivazione di meccanismi sempre più specializzati che ‘bloccano’ l’espressione specifica di determinati geni. Questo meccanismo, definito ‘deterministico’, è quello maggiormente utilizzato durante la specializzazione cellulare nel nostro organismo. Potremmo ipotizzare un meccanismo simile per le ORC in cui dalla combinazione di alcuni fattori trascrizionali si determina l’attivazione dell’espressione di un singolo OR.
Verifica della Teoria Deterministica
 Secondo questa teoria la combinazione casuale o pseudocasuale di fattori trascrizionali per esprimere un singolo OR non può essere disturbata dalla coespressione di un OR transgenico scelto da noi. Per verificare questa teoria ‘deterministica‘, quindi, possiamo generare un topo transgenico in cui alcune ORC esprimono in maniera forzata un determinato OR scelto da noi e valutare se c’è coespressione.
Secondo questa teoria la combinazione casuale o pseudocasuale di fattori trascrizionali per esprimere un singolo OR non può essere disturbata dalla coespressione di un OR transgenico scelto da noi. Per verificare questa teoria ‘deterministica‘, quindi, possiamo generare un topo transgenico in cui alcune ORC esprimono in maniera forzata un determinato OR scelto da noi e valutare se c’è coespressione.
Il risultato è interessante, non c’è alcuna sovrapposizione di segnali tra il OR espresso forzatamente da noi ed un qualsiasi altro recettore olfattivo endogeno. In pratica l’espressione forzata del nostro OR transgenico non permette di esprimere un qualsiasi altro OR endogeno, e se si tenta di generare un topo transgenico con una espressione forzata di due OR diversi nella stessa ORC, queste muoiono per apoptosi. Questi dati non sono di certo a supporto per la teoria deterministica, evidentemente la scelta del recettore olfattivo è un meccanismo molto più complesso. Ad aggiungere benzina sul fuoco c’è un’altra prova interessante, ovvero le ORC possono cambiare più volte il recettore odoroso espresso prima di prendere contatto con il cervello (maturazione cellulare). La teoria deterministica è molto fallace con questi risultati, poco probabile che sia quella buona.
La Teoria Stocastica
 Un’altra teoria che si è fatta strada nel tempo è basata sulla casualità della scelta del OR espresso (teoria stocastica). Per capirci immaginiamo un fattore trascrizionale unico che sia in grado di legare il promotore di tutti i geni dei OR e di consentirne l’espressione; tecnicamente può legarsi ad un solo OR e quindi può esprimere solo quello. Se il fattore trascrizionale non è molto efficiente nell’esprimere un OR si potrebbe spiegare come mai una ORC cambia OR nei primi stadi pur seguendo sempre la regola di esprimere un solo OR alla volta. In seguito, quando un OR espresso dà un forte segnale di funzione (feed-back) il fattore trascrizionale si fissa sul gene che sta trascrivendo fino allo stadio di maturazione.
Un’altra teoria che si è fatta strada nel tempo è basata sulla casualità della scelta del OR espresso (teoria stocastica). Per capirci immaginiamo un fattore trascrizionale unico che sia in grado di legare il promotore di tutti i geni dei OR e di consentirne l’espressione; tecnicamente può legarsi ad un solo OR e quindi può esprimere solo quello. Se il fattore trascrizionale non è molto efficiente nell’esprimere un OR si potrebbe spiegare come mai una ORC cambia OR nei primi stadi pur seguendo sempre la regola di esprimere un solo OR alla volta. In seguito, quando un OR espresso dà un forte segnale di funzione (feed-back) il fattore trascrizionale si fissa sul gene che sta trascrivendo fino allo stadio di maturazione.
E’ una teoria che funziona e che spiega anche i risultati dei topi transgenici; il OR transgenico blocca l’espressione di altri recettori attraverso un feed-back abberrante. Però, come al solito la teoria va convalidata, e c’è l’imbarazzo della scelta sul come; una prova schiacciante potrebbe derivare dall’eliminazione del feed-back recettoriale. In particolare potremmo alterare un gene OR in modo tale da codificare un recettore non funzionante pur conservandone la struttura genica.
I risultati dimostrano che il topo transgenico non esprime mai un OR non funzionante, probabilmente proprio per la mancanza del feed-back del recettore maturo necessario per fissare la scelta. Per esserne sicuri, è stata effettuata una controprova per dimostrare che le ORC tentano di esprimere il OR non funzionante, e che solo successivamente cambiano la scelta su un altro gene OR che funzioni. Per dimostrarlo si è generato un topo transgenico in cui l’espressione anche transiente del recettore non funzionante provoca un danno irreversibile al DNA della stessa cellula (tecnica Cre/LoxP), ed i risultati hanno mostrato che le ORC scelgono il OR transgenico e non funzionante con la stessa probabilità di tutti gli altri OR, sebbene questa scelta sia cambiata prima della maturazione cellulare per mancanza di feed-back. Un’altra prova conclusiva e schiacciante è che la generazione di un topo trangenico che esprime un OR esogeno e non funzionante non disturba l’espressione dei OR endogeni. Questo dimostra che è la funzione di un OR a bloccare l’espressione dei OR endogeni, non la struttura e l’espressione dei geni OR endogeni.
La spina nel fianco per la Teoria Stocastica
La teoria stocastica fino ad ora ha sempre soddisfatto tutti i dati sperimentali ottenuti, ma ciononostante ha un punto debolissimo. Questa teoria è retta dalla possibilità di poter innescare la trascrizione di qualsiasi dei 1.000 geni dei OR da uno o pochi fattori trascrizionali, e questo sarebbe possibile solo nel caso in cui i geni OR presentino una sequenza consenso che li identifichi nel genoma. Ebbene, nonostante molti ricercatori si siano impegnati nell’analisi di omologie di sequenze tra i geni degli OR, i risultati ottenuti sono stati alquanto deludenti; con i dati attuali si potrebbero raggruppare non più di una decina di geni degli OR per volta tirando in ballo meccanismi di riconoscimento improbabili.
E se fossero dei Linfociti?
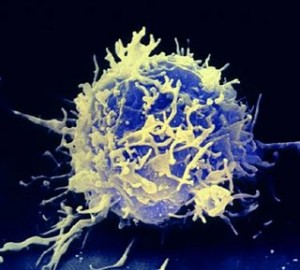 Se ci pensate, esiste un meccanismo di scelta molto simile che avviene nei linfociti. Queste cellule, però, non decidono l’anticorpo da esprimere in base a fattori trascrizionali, ma attraverso una ricombinazione genetica guidata. Si tratta di una piccola deviazione della teoria stocastica ed è anche risolutiva.
Se ci pensate, esiste un meccanismo di scelta molto simile che avviene nei linfociti. Queste cellule, però, non decidono l’anticorpo da esprimere in base a fattori trascrizionali, ma attraverso una ricombinazione genetica guidata. Si tratta di una piccola deviazione della teoria stocastica ed è anche risolutiva.
Come al solito dobbiamo convalidare tale teoria… ed escludiamo subito la proposta di sequenziare l’intero genoma di singole cellule, sarebbe difficilissimo e ci vorrebbero tanti anni! Possiamo ricorrere ad un trucco chiamato FISH (fluorescent in situ hybridization), ovvero rendere fluorescente una sonda ad RNA verso un introne di un OR. In questa tecnica non è possibile valutare una ricombinazione genica in maniera diretta, tuttavia è possibile localizzare dove inizia la trascrizione di un determinato gene all’interno del nucleo e valutare se, in seguito ad una ricombinazione, cambia posizione. I risultati, alquanto discutibili, dimostrano che il sito di inizio della trascrizione avviene sempre nello stesso punto del nucleo e del genoma, quindi niente ricombinazione, ma c’è una novità inaspettata; la trascrizione di un OR proviene solamente da un singolo allele.
Sbrogliare la matassa di informazioni non è mai facile

Spesso nella ricerca è importante definire delle ipotesi e porsi le domande giuste, ovvero cercare di identificare la cosa più importante di un meccanismo e trarne quante più informazioni possibili per verifica e deduzione. Nel nostro caso la codifica di un singolo allele, cioè solo 1 delle due copie genetiche di un OR, è ancora in linea con la teoria della ricombinazione genetica, poiché se è vero che la scelta è casuale, questa non può avvenire in entrambi gli alleli fratelli allo stesso modo. Inoltre se l’inizio della trascrizione avviene sempre nello stesso punto, questo non esclude che tutto il resto del DNA (es il promotore) possa essere stato ricombinato.
L’ipotesi che ne scaturisce è questa: La cellula sceglie un OR da esprimere in modo casuale, in mancanza di un feed-back funzionale cambia scelta, in seguito, alla maturazione cellulare la scelta diventa definitiva mediante ricombinazione di un promotore o di segnali a monte dell’OR scelto. Questa ipotesi potrebbe spiegare come mai l’espressione forzata di un recettore esogeno possa bloccare l’espressione dei recettori endogeni; probabilmente la proteina inserita provoca un feed-back che innesca una ricombinazione abberrante del DNA nei OR endogeni. La teoria per ora tiene ma con le parole non si dimostra mai niente, è fondamentale convalidare la teoria e questa volta non è per nulla facile evidenziare tante piccole ricombinazioni in singole cellule di 20 micron di diametro circa.
Come dimostrare una ricombinazione che si possa trovare in un qualsiasi punto del genoma? In questo caso il test di validazione è non semplicissimo. L’idea è che la ricombinazione del DNA in una ORC provocherebbe il blocco dell’espressione di un determinato OR in tutte le condizioni, come avviene per un linfocita. Se da questo DNA ricombinato si clonasse il topo stesso da cui deriva… l’animale che risulterebbe dovrebbe esprimere solo un OR, poiché il suo DNA è già ricombinato in maniera irreversibile. I risultati sono stati scioccanti, il topo che ne risulta è perfettamente sano e ha tutto l’apparato olfattivo intatto e funzionante come in un qualsiasi wild-type. Nonostante l’analisi di circa 100 kb a monte ed a valle del gene incriminato non c’è stata nessuna modifica genetica o epigenetica.
Basta un soffio ed il castello crolla
 Certamente la ricombinazione è da scartare ma fino ad ora niente ha smentito la casualità della scelta del OR. Tuttavia un gruppo di ricerca ha recentemente messo in discussione anche questa ipotesi granitica.
Certamente la ricombinazione è da scartare ma fino ad ora niente ha smentito la casualità della scelta del OR. Tuttavia un gruppo di ricerca ha recentemente messo in discussione anche questa ipotesi granitica.
L’idea di fondo è la seguente… Se è vero che la cellula sceglie casualmente un gene su circa 1.000, e di questo solo 1 allele, ne consegue che la scelta è di 1 sui 2.000 alleli disponibili. Se riuscissimo ad inattivare solo 1 di questi 2.000 alleli, ad esempio in modo tale che esprima irreversibilmente EGFP (tecnica Cre/loxP), potremmo rendere fluorescenti le cellule che hanno ‘cambiato idea’ per mancanza di feed-back e verificare quale altro gene sceglie dei 1.999 rimanenti.
Il risultato, ancora una volta, è assolutamente strabiliante. Il topo transgenico presenta una coespressione di EGFP solo con il suo allele funzionante corrispondente. In poche parole l’espressione di EGFP, come previsto, non dà feed-back positivo per cui la cellula cambia OR da esprimere; ma la seconda scelta non è per niente casuale, qualcosa guida la scelta sull’allele di ‘backup’ presente sul cromosoma fratello e non sugli altri 1.998 alleli rimanenti. Come potrebbe la cellula riconoscere in modo specifico l’altro allele se la scelta è basata solo sulla casualità? Qualcosa non quadra, ed in questo modo la casualità che ha retto la teoria stocastica fino ad ora è stata, almeno parzialmente, distrutta in un soffio.
L’olfatto che guida l’assone durante il lungo cammino
 L’ultima domanda è ‘Cosa succede con il collegamento al cervello quando forziamo una cellula ORC ad esprimere un recettore specifico scelto da noi?’. Gli esperimenti mostrano che il collegamento cervello-recettore è perfetto, quindi ne consegue che il collegamento è basato solo sull’OR espresso.
L’ultima domanda è ‘Cosa succede con il collegamento al cervello quando forziamo una cellula ORC ad esprimere un recettore specifico scelto da noi?’. Gli esperimenti mostrano che il collegamento cervello-recettore è perfetto, quindi ne consegue che il collegamento è basato solo sull’OR espresso.
Ma quando questa scelta cambia all’improvviso?
Per rispondere a questa domanda un gruppo di ricercatori ha tentato di cambiare il recettore scelto dalla cellula poco prima che questa riesca ad ‘agganciarsi’ al cervello (maturare), attraverso un gene OR esogeno ed inducibile.
In questo topo transgenico le ORC scelgono un recettore endogeno in un modo apparentemente casuale, e cominciano la propagazione dell’assone verso il cervello cercando la zona giusta per formare sinapsi. Durante questa fase si può innescare l’espressione del recettore transgenico con il conseguentemente spegnimento del recettore endogeno…
La cellula ancora immatura, come abbiamo visto, si ‘adatta’ al nuovo recettore, ma si aggancerà anche alla zona cerebrale giusta? La risposta è incredibilmente sì. il collegamento e la funzione del recettore è perfetto nonostante il cambio recettoriale durante la formazione dell’assone. Tuttavia, se si cerca di cambiare la scelta dell’OR espresso dopo la maturazione della ORC, si provoca una abberrazione che innesca apoptosi.
Come è possibile che un OR possa guidare la scelta della regione cerebrale giusta a cui ‘agganciarsi’ senza avere altri 1.000 geni deputati al riconoscimento dei diversi OR da parte del cervello?
Qualcuno ha suggerito un’ipotesi alquanto affascinante, secondo cui un OR si trovi alla testa dell’assone durante la fase di elongazione, guidandolo verso il corretto punto di aggancio al cervello come farebbe un cane da tartufo a portarci verso questo prezioso tubero in un intero bosco.
Teoria affascinante e stravagante, tuttavia ancora insoluta ed in cerca di qualcuno che la convalidi o la controbatta con esperimenti, o con ipotesi più semplici.
Conclusione
In questo lungo post abbiamo messo sul piatto tutto quello che è noto sull’olfatto senza uscirne fuori con una teoria solida in grado di spiegare tutti i dati sperimentali. Per chi fa ricerca questo risultato è il pane quotidiano poiché non si seguono mai strade battute, si cerca sempre di crearne nuove, con nuove ipotesi, nuove verifiche e nuove deduzioni che porteranno ad altre ipotesi che si spera siano migliori delle precedenti e più vicine alla verità.
Tutt’ora ci sono gruppi di ricerca che analizzano le seguenze di DNA dei recettori OR cercando qualcosa in comune; analisti in cerca di modifiche epigenetiche in questi clusters genomici; cacciatori di nuovi meccanismi cellulari; esperti di transgeni che tentano nuove e più affascinanti teorie, a volte anche bizzarre… e poi ci sono ricercatori che provano a dare un senso a questo puzzle di dati disarmonici. Il tutto per svelare un mistero che dura da tanti anni oramai.
Non vi è dubbio che ci sono persone che provano un certo fascino nel partecipare a questa continua caccia alla soluzione del problema strato dopo strato, tirando fuori teorie sempre più complesse ed affascinanti, ed a volte come in questo caso si tirano fuori addirittura nuovi meccanismi che si ignoravano fino a poco tempo fa.
Un in bocca al lupo a chi si addentrerà nel risolvere questo mistero anche solo per 5 minuti ed un grazie a Nico e Patrizio per l’incoraggiamento ed i consigli che mi hanno dato per la stesura di questo post.
Tags:
Animali transgenici,
Apoptosi,
Biologia molecolare,
Cellule Recettoriali,
Cre/LoxP,
fattori trascrizionali,
mappa sensoriale,
Neuroni,
neuroni olfattivi,
Olfatto,
Percezione
15 maggio 2009 - 12:03 pm
Rieccoci ancora per un’altra puntata per la divulgazione scientifica sull’ischemia cerebrale. Questo che vedremo è, in ordine cronologico, l’ultimo capitolo da me scritto ma il primo da me adattato per la pubblicazione di questa serie. Il motivo è che, a parer mio, potrebbe essere più intuitivo per le persone che non si sono mai affacciati ad un discorso scientifico impegnato. Trattasi di circa 15 anni di ricerche in questo campo seguito e presentato per lo più in ordine cronologico con qualche inesattezza o approssimazione in qualche punto per rendere il discorso quanto più scorrevole e divulgativo possibile; Vogliate scusarmi se in qualche caso ho semplificato un po’ troppo quanto si conosce realmente in letteratura.
Preambolo
Per chi non avesse seguito il percorso del nostro misterioso ‘giallo’ sull’ischemia cerebrale forse sarebbe bene leggersi i capitoli precedenti per poter seguire meglio questo ‘racconto’
http://www.molecularlab.it/insideneuroscience/?p=26
http://www.molecularlab.it/insideneuroscience/?p=37
http://www.molecularlab.it/insideneuroscience/?p=41
Siamo nelle vesti di un improbabile investigatore molecolare con il compito di risolvere un misterioso omicidio di cellule neuronali in seguito ad ictus. Fino ad ora abbiamo cercato di ricostruire la vita dei neuroni, le ultime ore delle vittime e come si presentano agli occhi della scena del crimine. In questo capitolo, invece, cercheremo di puntare il dito sul primo imputato eccellente dell’ischemia, ovvero un fattore trascrizionale ‘particolare’. Già la sola appartenenza di una proteina alla categoria “fattore trascrizionale” conferisce quell’alone di mistero ed ambiguità tipico, infatti, è noto che queste proteine frequentano “strane compagnie”, tale che il più amabile difensore della vitalità cellulare può trasformarsi nel peggior nemico da abbattere nelle neoplasie.
Ecco a voi quanto segue.
La prima pista delle indagini
 Una delle osservazioni più ovvie che mi sono state fatte nel tempo si basa sul fatto che tutto quello che si avvia durante l’ischemia cerebrale è senza alcun dubbio innescato dalla mancanza di ossigeno e nutrimento… secondo questa ipotesi ci si dovrebbe concentrare sugli eventi che sono attivati dall’ipossia e/o glucopenia per arrivare al vero meccanismo che innesca la morte neuronale ritardata. L’osservazione è ottima, e tantissimi ricercatori si sono concentrati su questa pista per trovare i responsabili.
Una delle osservazioni più ovvie che mi sono state fatte nel tempo si basa sul fatto che tutto quello che si avvia durante l’ischemia cerebrale è senza alcun dubbio innescato dalla mancanza di ossigeno e nutrimento… secondo questa ipotesi ci si dovrebbe concentrare sugli eventi che sono attivati dall’ipossia e/o glucopenia per arrivare al vero meccanismo che innesca la morte neuronale ritardata. L’osservazione è ottima, e tantissimi ricercatori si sono concentrati su questa pista per trovare i responsabili.
Sicuramente uno dei target più dibattuti in questo contesto è il fattore trascrizionale che porta il nome di Hypoxia Inducible Factor (HIF), ovvero fattore indotto dall’ipossia. Ce ne sono 3, chiamati HIF-1, HIF-2 e HIF-3; di questi solo il primo è noto alla maggioranza della comunità scientifica, e sembra essere il più importante.
Quale personaggio per HIF-1?
 Se dovessi associare questo fattore trascrizionale ad un personaggio di un giallo, potrebbe essere rappresentato senza alcun dubbio da un antipatico ed ambiguo individuo che attira su di sé tutti gli indizi, in modo tale che rappresenti l’assassino nell’immaginario del lettore per buona parte del racconto. In genere queste persone durante il racconto possono trovarsi alternativamente tra i principali sospetti, tra i fiancheggiatori, tra gli innocenti o tra i più improbabili difensori della verità.
Se dovessi associare questo fattore trascrizionale ad un personaggio di un giallo, potrebbe essere rappresentato senza alcun dubbio da un antipatico ed ambiguo individuo che attira su di sé tutti gli indizi, in modo tale che rappresenti l’assassino nell’immaginario del lettore per buona parte del racconto. In genere queste persone durante il racconto possono trovarsi alternativamente tra i principali sospetti, tra i fiancheggiatori, tra gli innocenti o tra i più improbabili difensori della verità.
Assassino, complice, arma, ancora di salvezza e persino futile perdita di energia… queste sono le definizioni che nel tempo sono state associate a questo strano fattore trascrizionale.
I Primi indizi ed il primo incontro
La pista dell’ipossia ci porta a chiedere se la scena del crimine era frequentata da fattori che sono proprio regolati da questa condizione. Da un elenco di proteine potremmo scorgere un personaggio molto influente definito HIF-1 e già qui giungiamo al primo mistero; dall’analisi della zona ischemica emerge chiaramente che HIF-1 non è presente al momento del ‘delitto neuronale’, un semplice Western Blot per l’analisi delle proteine mostra chiaramente una debolissima banda di HIF-1 ma niente di più di quello che si troverebbe in un qualsiasi tessuto in condizioni normali. Il personaggio quindi sembra non avere indizi a carico, tuttavia ci sono delle stranezze da puntualizzare.
Tiriamo fuori il nostro asso nella manica, ovvero la ricostruzione dell’avvenimento in un modello di ischemia su animale ed analizziamo la quantità di HIF-1 in diverse aree cerebrali a diversi tempi dopo l’occlusione dell’arteria cerebrale media. Dai risultati ci renderemo conto che HIF-1 non era presente prima dell’occlusione e neanche all’inizio dell’occlusione stessa, mentre è presente per pochissimo tempo durante l’ipossia neuronale per poi sparire di nuovo improvvisamente subito dopo il ripristino del circolo sanguigno.
HIF-1 dovrebbe spiegarci molte cose al riguardo sul perché c’era, cosa ha fatto e come ha fatto ad apparire e scomparire velocemente in circostanze quantomeno sospette.
Il mago Houdinì – Apparizioni e Sparizioni
HIF-1 è un fattore trascrizionale composto da due sub-unità definite HIF-1alfa e HIF-1beta, prodotti da due geni diversi molto conservati nell’evoluzione dei mammiferi. La trascrizione e la formazione di queste due proteine sono costanti ed ubiquitari in tutto l’organismo, tuttavia con un banale western blot non è possibile vedere una buona dose di proteina nei tessuti.
Il perché è subito chiarito dal suo scopritore Gregg Semenza, che ha dimostrato come in condizioni di ossigenazione ‘normale’ HIF-1alfa è idrossilato attraverso un enzima e poi processato per la degradazione… in poche parole HIF-1alfa è continuamente prodotto ma è anche degradato subito dopo in presenza di ossigeno molecolare. Infatti, quando l’ossigeno comincia a mancare HIF-1alfa non si può più degradare e si accumula legandosi al suo cofattore HIF-1beta formando il fattore trascrizionale completo HIF-1 che va ad attivare una serie di geni target. In seguito alla riossigenazione HIF-1alfa è nuovamente degradato ed il complesso HIF-1 non si forma più.
Ecco così spiegato il mistero della sparizione ed apparizione di HIF-1 durante l’ischemia cerebrale.
Conosci gli amici che frequenti e saprai chi sei
La domanda più interessante a questo punto è “quali geni target attiva HIF-1?”. Bhé ce ne sono centinaia, e sono divisi in classi secondo il meccanismo globale a cui afferiscono.
Troviamo geni del metabolismo anaerobico del glucosio, ovvero quegli enzimi che servono per ottenere energia dal glucosio senza la necessità di ossigeno, troviamo anche geni che servono per aumentare l’ossigenazione in una certa regione del tessuto come ad esempio VEGF il fattore che induce la vascolarizzazione in loco per apportare più ossigeno e nutrienti. Tra i target c’è anche l’eritropoietina che aumenta la produzione di globuli rossi che portano l’ossigeno in tutto l’organismo.
Poi ci sono geni un po’ più strani che mirano comunque alla sopravvivenza della cellula in condizioni anaerobiche e geni di cui non si sa bene la funzione nell’ipossia.
Tutto sommato è un ‘buono’ e serve alle cellule per ‘adattarsi’ alle scarse riserve di ossigeno e soprattutto promuovere il rimodellamento dell’organismo per avere più ossigeno.
Buono?
C’è anche l’altra faccia della medaglia, come per ogni fattore trascrizionale. L’altra faccia della ‘Janus Face’ HIF-1 è che se passa ‘dall’altra parte’ favorisce la vascolarizzazione di neoplasie (tumori) rendendolo più maligno e più resistente alle terapie antineoplastiche. Non a caso le mutazioni dei geni che servono per degradare HIF-1 in presenza di ossigeno siano dei noti oncogeni, ovvero geni cruciali che favoriscono la formazione e l’accrescimento dei tumori quando non funzionano a dovere.
Cosa ci faceva HIF-1 durante l’ischemia?
 Bene dopo questa velocissima presentazione di questo angelo o diavolo è ora di chiedersi cosa stava facendo subito dopo l’occlusione dell’arteria cerebrale e proprio in quei neuroni che moriranno diverse ore dopo… Stava avvelenando il loro futuro o stava costituendo un disperato tentativo di salvataggio?
Bene dopo questa velocissima presentazione di questo angelo o diavolo è ora di chiedersi cosa stava facendo subito dopo l’occlusione dell’arteria cerebrale e proprio in quei neuroni che moriranno diverse ore dopo… Stava avvelenando il loro futuro o stava costituendo un disperato tentativo di salvataggio?
Se provate a chiedere a HIF-1 cosa stesse facendo la risposta è ovvia, stava promovendo la sintesi di geni del metabolismo anaerobico per consentire ai neuroni di far fronte all’immediata assenza di ossigeno.
Facendo una Cromatin Immunoprecipitation (ChIP), ovvero una tecnica che ci permette di determinare se un fattore trascrizionale è legato o no ad una determinata regione genomica, c’è un primo riscontro positivo, effettivamente HIF-1 si trovava nelle posizioni di attivazione dei geni che servono per il metabolismo anaerobico.
Però c’è un’incongruenza, il tempo necessario ad HIF-1 per attivare i geni del metabolismo è costituito almeno da diverse ore; ovvero il tempo necessario per aumentare la sintesi di mRNA e di proteine. HIF-1, invece, rimane legato ai geni descritti per pochissimo tempo, al massimo un’ora o anche molto meno, un tempo che non basterebbe neanche a reclutare tutti i cofattori necessari per l’attivazione della trascrizione genica… come mai?
HIF-1 si difende esponendo i propri fatti… ovvero si trovava lì al momento dell’ipossia, aveva visto che ‘la nave stava affondando nell’ipossia’ e quindi si è attivato quanto prima come da suo dovere, poi è subentrata la riossigenazione dell’area cerebrale che lui non poteva prevedere ed è stato degradato dall’ossigeno prima che potesse fare effettivamente qualcosa. Per cui in realtà si tratterebbe di un tentativo andato a vuoto.
Dall’analisi per RT-PCR, tecnica che permette di determinare l’aumento o la diminuzione dell’espressione di determinati geni, c’è una prima conferma dell’alibi di HIF-1. Effettivamente stavano aumentando, seppur di poco, la sintesi dei geni implicati nel metabolismo anaerobico, del tutto sovrapponibile con quello che ci si aspetterebbe dall’azione esplicata da HIF-1.

Innocente allora?
Prima di prosciogliere l’imputato io suggerirei di analizzare anche chi ha frequentato HIF-1 in quella notte misteriosa dove è avvenuta l’ipossia. C’erano altri geni che ha indotto HIF-1?
Un gene molto importante è VEGF come già accennato, implicato nell’induzione della vascolarizzazione a lungo termine, ma che ha anche un’azione neuroprotettiva acuta nei confronti dell’ischemia cerebrale. Poi ci sono l’eritropoietina, che serve per aumentare i globuli rossi nel sangue, anch’esso neuroprotettivo in fase acuta, e tanti altri geni più o meno noti per avere un’azione neuroprotettiva. Un eroe dunque.

Il lato oscuro di HIF-1
C’è però anche chi fa notare che i fattori trascrizionali in condizioni non fisiologiche come l’ischemia cerebrale possono frequentare anche ‘compagnie diverse’ ed insolite da quelle catalogate. Infatti, da alcuni esperimenti di Microarray, tecnica che permette di vedere l’aumento o la diminuzione dell’espressione di un ampio numero di geni in condizioni fisiologiche rispetto a quelle patologiche, è emerso che HIF-1 ha scarsamente indotto geni neuroprotettivi, come già descritto, però durante la sua attivazione sono aumentati enormemente anche tanti geni di ‘malaffare’.
Insomma tanto basta per iniziare il processo contro questo ‘strano individuo’
Che il processo abbia inizio… entri il primo imputato HIF-1
 La parola all’accusa
La parola all’accusa
Quei pochi minuti che bastano
Sebbene siano necessarie diverse ore per attivare i geni del metabolismo anaerobico, per HIF-1 sono stati necessari pochi minuti per attivare alcuni dei geni più distruttivi dell’ischemia. E non si tratta di una piccolo aumento di espressione. Il gene in questione è la nitrossido sintetasi inducibile che produce radicali liberi sottoforma di gas nitrossido con un’azione neurotossica letale per i neuroni. Da un’analisi con Decoy, ovvero la somministrazione di piccoli tratti di DNA a doppio filamento che simulano il sito di legame di HIF-1 e lo sequestrano lontano dai geni bersaglio, è stato possibile determinare con certezza che HIF-1 è il principale responsabile dell’aumento della trascrizione della iNOS. Quando ad una cellula ipossica si dà il Decoy, HIF-1 lo lega e non riesce a raggiungere il gene bersaglio, un effetto simile ad un knock-out genico. La iNOS è nota per causare la morte cellulare dei neuroni, una questione indubbia.
Ma come è possibile che ad HIF-1 bastino pochi minuti per l’attivazione di iNOS mentre sono necessarie ore per i geni neuroprotettivi? Greeg Semenza è riuscito a chiarire anche questo altro aspetto. HIF-1, infatti, è prodotto continuamente dalla cellula, ed è anche degradato continuamente, ma una piccola quota di HIF-1 è presente nelle cellule, visibile per western blot, e basta per legare ed indurre determinati geni insoliti. Talvolta li lega anche quando non c’è ipossia ed attende la mancanza di ossigeno così per trovarsi già in loco quando serve.
Insomma HIF-1 è stato visto aumentare e fomentare geni per la degenerazione neuronale, mentre i geni neuroprotettivi non erano stati trattati allo stesso modo, dando solo un’apparenza di eroe salvatore.
A carico di HIF-1 c’è anche la precisione di espressione proprio nei neuroni e nei luoghi dove ci sarà, diverse ore la sua scomparsa, la maggior percentuale di neuroni morti. Una coincidenza forse?
Insomma, la situazione si è messa male per HIF-1, le prove vanno decisamente contro; il legame degli enzimi metabolici e neuroprotettivi potrebbe essere solo una copertura per il vero scopo, ovvero la degenerazione dei neuroni?
La parola alla difesa
 La difesa tira fuori il dogma scientifico
La difesa tira fuori il dogma scientifico
Il Dogma scientifico
Per chi non lo sapesse, esiste un dogma per ogni disciplina culturale umana e la biologia non ne è immune. Il Dogma dei biologi è che niente succede a caso, poiché per ogni gene o proteina ci sono milioni di anni di evoluzione che raffina tutti i meccanismi a quelli fondamentali per la sopravvivenza e per l’evoluzione. Se ci fosse qualcosa di inutile o peggiorativo sarebbe eliminato automaticamente dall’evoluzione stessa. Per cui la selezione e la conservazione di un fattore indotto dall’ipossia che causi morte nei neuroni non è credibile poiché non darebbe un vantaggio per la sopravvivenza.
Dopotutto quello che si vede è una sommatoria di tantissimi meccanismi che interagiscono tra di loro e quello che si osserva è solo la summa del tutto da cui è difficile estrapolare un singolo messaggio. Meglio ricorrere all’incidente probatorio…
Ovvero spegniamo HIF-1 con un knock-out genico ed induciamo l’ischemia, verifichiamo in questo caso se l’animale che manca di questo fattore è più o meno esposto al danno neuronale rispetto agli animali wild-type.
L’incidente probatorio o la prova del nove
La prova più importante seguita da quella definitiva molto spesso è la generazione di un topo knock-out per il gene di interesse. Ci sono stati tantissimi esempi di questo tipo, vale sia come inizio di una ricerca per capire grossolanamente un gene di cosa sia responsabile e vale anche come prova raffinata per avere la prova definitiva che un determinato gene sia coinvolto in un certo meccanismo.
Purtroppo nonostante intensi sforzi per ottenere topi knock-out per HIF-1alfa, è emerso che un animale privato di questo gene non riesce a svilupparsi ed a sopravvivere ai primi stadi embrionali, anche se è esposto ad un normale livello di ossigenazione. Ovviamente questo è stato il motivo dei ritardi nel fare un topo transgenico e quindi definire una prova evidente del suo ruolo in tanti processi fisiologici e patologici.
Tuttavia, recentemente è stato possibile generare un topo con un knock-out tessuto specifico di HIF-1alfa per i neuroni, il risultato è stato quantomeno sorprendente… l’assenza di HIF-1alfa durante l’ischemia cerebrale ha enormemente potenziato i danni neuronali.
Un fulmine a ciel sereno per tutti… L’imputato è senza dubbio da scagionare e da mettere tra i difensori cellulari.
Conclusioni
In questo capitolo abbiamo visto un esempio di imputato eccellente dell’ischemia cerebrale che ancora oggi per alcuni gruppi di ricerca rappresenta ancora il centro dell’attenzione. Gli studi andranno sicuramente avanti, anche se non ci si aspetterebbe ancora molto da quanto descritto, HIF-1alfa nel processo dell’ischemia cerebrale si potrebbe definire almeno non pericoloso tanto quanto altri imputati eccellenti che vedremo nelle prossime puntate. Per questi motivi l’attenzione su questo fattore trascrizionale si è concentrato soprattutto sulle neoplasie, ed ha tralasciato progressivamente l’ischemia cerebrale, renale e cardiaca. Vedremo se qualcuno in futuro troverà qualche prova rilevante per riaprire nuovamente il processo. In attesa di tale data ci rivedremo nella prossima puntata dove vedremo un altro imputato importante.
Tags: fattori trascrizionali, HIF-1, HIF-1alfa, Ictus, Ischemia Cerebrale, Mente, Neuroscienze
7 gennaio 2008 - 3:20 pm
Uno dei motivi per cui leggo spesso le riviste con altissimo impact factor è la speranza di trovare un articolo davvero rivoluzionario che possa chiarire un determiato meccanismo oppure possa aprire la strada verso nuove prospettive, oltre che per aggiornarmi sulle recenti scoperte. Purtroppo devo dire che le riviste scientifiche più importanti alla fine non pubblicano sempre cose drasticamente innovative o chiarificatrici, anzi a volte le riviste un po’ più modeste contengono articoli con informazioni molto più convincenti e serie rispetto a Nature, Cell, Neuron etc.
In una di queste letture, mi è capitato di leggere qualcosa su Cell che, a mio avviso, è veramente incredibile, oltre che curioso. L’articolo in questione tratta di un canale del calcio voltaggio dipendente (Cav1.2) che ha anche un secondo ruolo funzionale del tutto diverso ed imprevedibile da quello che ci si aspetterebbe da un semplice canale.
Prima di entrare nel merito bisognerebbe introdurre il concetto di canale voltaggio dipendente oramai acquisito da tanti anni nel campo delle neuroscienze ma che per i non addetti ai lavori potrebbe sembrare fuorviante.
Diciamo che almeno qualche migliaio dei circa 30.000 geni umani codificano per proteine di membrana che possono formare dei canali idrofilici attraverso cui, in particolari condizioni, possono passare degli ioni. Il motivo per cui esistono tante proteine non è dovuto alla presenza tanti tipi di ioni da far passare, ma dalla fine regolazione a cui questi canali devono rispondere. La quasi totalità di questi canali ionici sono sempre chiusi, ma possono aprirsi in determinate condizioni secondo una cinetica caratteristica che è dipendente dai componenti da cui è formato.
Alcuni canali possono aprirsi solamente in seguito al legame di un neurotrasmettitore sulla regione esterna, o interna, del complesso proteico (Recettori canale), come avvine per il recettore nicotinico che fa entrare sodio all’interno della cellula in seguito al legame dell’acetilcolina con la parte esterna del recettore; la cinetica di apertura/chiusura o l’affinità con l’agonista recettoriale dipende dai componenti del canale stesso. Ovviamente l’ingresso di sodio che ne scaturisce provoca a sua volta l’attivazione di una serie di altri meccanismi a cascata come le tessere del domino che portano all’attivazione di altri effettori. Altri canali possono invece aprirsi in seguito al cambiamento delle condizioni cellulari, variazione di pH (es ASICs), di concentrazioni ioniche (es Connessine) oppure del potenziale di membrana (es canali voltaggio dipendenti).
Ogni tipo cellulare possiede il proprio corredo di recettori canale, canali voltaggio dipendenti, canali passivi e trasportatori; l’insieme di tali complessi proteici con caratteristiche finemente diverse tra loro determina delle differenze macroscopiche e microscopiche nella reattività cellulare, forma del potenziale d’azione, velocità di risposta ad uno stimolo, eccitabilità di membrana, potenziale di membrana a riposo, picco di depolarizzazione e tantissime altre variabili che fanno di un neurone un sottotipo cellulare unico. Non entro nel dettaglio di quanti canali del calcio voltaggio dipendenti esistono, dico solo che sono identificate in varie classi con la sigla Cav seguita da un numero che identifica la classe seguito da un punto e poi un altro numero che identifica il sottotipo. Tutti questi canali regolano direttamente o indirettamente tante funzioni neuronali, come la memoria, il dolore, l’eccitabilità, la fertilità e la sensibilità a determinati danni patologici. Tuttavia è noto anche che questi canali ionici possono avere anche una funzione nella regolazione della trascrizione genica, poiché molti fattori trascrizionali sono regolati dalle concentrazioni di determinati ioni come ad esempio il calcio. Questa è stata sempre una teoria debole, poiché indubbiamente ci sono dei casi in cui l’effetto sulla trascrizione genica può essere drasticamente diverso se si trasfettano in cellule dei canali composti da subunità diverse. Insomma se è l’ingresso di ioni calcio a determinare l’attivazione di alcuni fattori trascrizionali allora l’effetto dovrebbe essere uguale per qualsiasi canale che fa entrare calcio, soprattutto se 4 subunità su 5 sono identiche e le caratteristiche cinetiche di ingresso e uscita del calcio sono praticamente le stesse.
Tuttavia questo non avviene sempre, come ho già accennato, l’effetto può essere drasticamente diverso, tale che alcuni canali ionici possono determinare il differenziamento neuronale in alcune cellule, mentre altri canali possono determinare l’espressione di proteine del metabolismo o neurotrasmettitori in maniera non chiara.
Una spiegazione che, secondo me, ha aperto la strada e gli occhi a molti ricercatori scientifici risiede nell’articolo di cui parlavo all’inizio di questa disquisizione, pubblicato su un numero di Cell dell’anno scorso dalla dott.sa Gomez-Ospina dell’università di Stenford. Il canale del calcio voltaggio dipendente (Cav1.2) possiede nel dominio C-terminale una sequenza peptidica che, se tagliata, è di fatto un fattore trascrizionale a tutti gli effetti e modula un cospicuo gruppo di geni endogeni importanti per la neurotrasmissione e per l’eccitabilità neuronale.
Il fattore trascrizionale in questione è stato definito Fattore Trascrizionale Associato al Canale del Calcio (CCAT) e si origina mediante la proteolisi di Cav1.2. Appena liberato, questo fattore trascrizionale si localizza nel nucleo dei soli neuroni grazie a meccanismi non ancora ben chiari e lì lega proteine nucleari e sequenze specifiche di DNA attivando la trascrizione dei geni target.
La cosa curiosa è che CCAT a sua volta è regolato dalle concentrazioni di calcio e dalla presenza di altri fattori trascrizionali che variano durante lo sviluppo, quindi il suo funzionamento è regolato da diverse variabili. L’effetto finale che ha il canale Cav1.2 quindi non è semplicemente variare la concentrazione di calcio ma regolare direttamente la trascrizione di geni target.
Bhé in definitiva questo articolo ci racconta come a volte le teorie che vengono formulate e che leggiamo di continuo non siano altro che un velo oltre il quale si nascondono dei meccanismi, come in questo caso, inimmaginabili e forse anche al limite della credibilità secondo le attuali conoscenze molecolari.
Forse non tutti i meccanismi sono di questo tipo e così semplici da spiegare, ma possiamo essere certi che c’è ancora tanto da scoprire dietro le nostre attuali teorie.
Tags:
Biologia molecolare,
Canali voltaggio dipendenti,
fattori trascrizionali,
Neuroscienze,
Potenziali d'azione
 Commenti disabilitati
Commenti disabilitati
