4 dicembre 2009 - 11:41 pm
Preambolo
Dopo aver preso in esame l’olfatto, ora vedremo un altra misteriosa percezione che mi sta molto a cuore; l’udito.
Sicuramente si tratta del più evoluto dei proverbiali 5 sensi, poiché é stata la fonte per le nostre intercomunicazioni sociali al più alto livello. Mediante la comunicazione abbiamo organizzato battute di caccia di gruppo, abbiamo chiesto aiuto, ci siamo accorti del pericolo e poi abbiamo cominciato a discutere di problemi, sentimenti fino ad arrivare alla poesia ed alla musica.
Il suono

Come al solito è necessario qualche punto di riferimento per metterci d’accordo e questa volta inizierò con la definizione di suono. Non vi è alcun dubbio che per suono si intenda una forma di energia cinetica (di tipo vibrazionale) che si propaga mediante un mezzo (aria, acqua, o altro) fino a raggiungere il nostro orecchio che ce lo rende percettibile attraverso il timpano e comprensibile attraverso connessioni nervose.
Il suono come energia

L’esempio più percettibile del suono inteso come propagazione di energia è quel fenomeno ben noto ai vecchi musicisti che accordavano il proprio strumento con il diapason. Il diapason, per chi non lo sapesse, è generalmente un pezzo di metallo a forma di Y che una volta colpito vibra con una particolarissima frequenza (440 Hz) chiamata ‘LA’ (in americano ‘A’). E’ utilizzato come suono di riferimento per accordare uno strumento (ad esempio la chitarra) sulla nota musicale LA, e poi da questa accordatura si possono ottenere tutte le altre note per trasposizione.
E’ esperienza comune che quando lo strumento è accordato bene, basta semplicemente avvicinare il diapason vibrante alle corde della chitarra per far vibrare magicamente la corda del LA senza toccarla. Quando ero giovane e suonavo la chitarra stupivo sempre mio cugino piccolo con questo fenomeno ben noto con il nome di risonanza. La spiegazione è semplice, l’energia cinetica del diapason riesce ad interagire in fase con la corda bersaglio che ne acquisisce l’energia, un po’ come sull’altalena, in cui c’è bisogno di dare la spinta con una certa frequenza per generare ed aumentare l’oscillazione del sellino. Questo fenomeno è valido a tutti i livelli tanto che il termine è stato poi traslato anche in altre discipline, come ad esempio nella chimica-fisica dove il termine di risonanza ed energia compatibile sono molto usati.
La musica nella scienza
 L’insieme strutturato ed armonico di suoni è senza alcun dubbio la musica e l’importanza di questa arte nella nostra specie non ha eguali in natura ed é anche correlata strettamente alle scienze. L’origine stessa della musica si perde nella preistoria ed è progredito parallelamente al nostro linguaggio fino ai tempi moderni. La perfetta correlazione tra la musica, la matematica, la scienza e l’arte non ha eguali.
L’insieme strutturato ed armonico di suoni è senza alcun dubbio la musica e l’importanza di questa arte nella nostra specie non ha eguali in natura ed é anche correlata strettamente alle scienze. L’origine stessa della musica si perde nella preistoria ed è progredito parallelamente al nostro linguaggio fino ai tempi moderni. La perfetta correlazione tra la musica, la matematica, la scienza e l’arte non ha eguali.
Non a caso il premio Nobel per la letteratura Hermann Hesse, nel suo mirabile capolavoro ‘il gioco delle perle di vetro‘ (1943) mette al centro dell’universo scientifico proprio la musica. Il romanzo di fantasia ambientato in un lontano futuro, infatti, parla di una tale frammentazione scientifica per le eccessive specializzazioni che non é più possibile una intercomunicazione tra i ricercatori. L’obiettivo del gioco delle perle di vetro é proprio quello di coniugare argomenti apparentemente tanto lontani in un unico concetto condiviso. Tra queste arti, proprio la musica é il principale capostipite, in quanto é già di fatto armonia di matematica, fisica, energia, ingegneria, arte, cultura, linguaggio e sentimento poetico in maniera semplice.
Le note come nomi
 Sappiamo tutti che a seconda della frequenza di vibrazione (Hertz) del suono si possono definire delle note raggruppate in periodi (ottave). L’insieme delle note stesse formeranno la musica la cui origine si perde nella preistoria.
Sappiamo tutti che a seconda della frequenza di vibrazione (Hertz) del suono si possono definire delle note raggruppate in periodi (ottave). L’insieme delle note stesse formeranno la musica la cui origine si perde nella preistoria.
Fin qui tutto già noto, ma pochi sanno che la musica ha avuto un ruolo sociale di intercomunicazione che è progredito parallelamente al linguaggio durante la nostra evoluzione, quindi ha quantomeno centinaia di migliaia di anni. Tuttavia, la formalità di questa arte, intesa come scrittura è recentissima, meno di un secolo.
Se prendiamo in esame i nomi ed il numero delle note, infatti, é solo un caso che si é concordati all’uso iniziale di 7 note periodiche a cui sono state aggiunte successivamente altre 5 (diesis o bemolle).
Anticamente si usava una notazione di origine greca che utilizzava le lettere dell’alfabeto α, β, γ etc per indicare le note. Tale notazione è ancora in uso nei paesi di lingua inglese con le lettere latine:
A = La, B = Si, C = Do, D = Re, E = Mi, F = Fa, G = Sol
Successivamente sappiamo che questa notazione è stata cambiata, tuttavia pochi sanno che il nome attuale delle note non é riferito al suono onomatopeico (tipo “bau” per l’abbaiare dei cani e “miao” per il miagolio dei gatti), bensì ad una convenzione stabilita arbitrariamente da Guido d’Arezzo nel XII secolo. Il nome delle note corrisponde al nome delle sillabe iniziali dei primi sei versetti dell’inno “Ut queant laxis”
« Ut queant laxis | Resonare fibris | Mira gestorum | Famuli tuorum | Solve polluti | Labii reatum, | Sancte Iohannes »
Traduzione
« Affinché i tuoi servi possano cantare con voci libere le meraviglie delle tue azioni, cancella il peccato, o santo Giovanni, dalle loro labbra indegne »
Nel XVI secolo la settima nota riceve il suo nome definitivo ‘Si’ e nel XVII secolo in Italia la nota ‘Ut’ viene sostituita con il nome attuale Do (da Dominus o da quello che l’ha proposto Giovanni Battista Doni) per la difficile pronuncia. In Francia questa modifica non venne fatta, e ancora oggi si usa il nome “ut” per la prima nota.
Cosa c’entra il suono con le Neuroscienze?
 Una volta, in un seminario del nostro dottorato ci venne a trovare un professore che provò a spiegare cosa fosse e dove si trovasse la nostra coscienza. Non ricordo nulla di interessante in quelle parole, ma una cosa mi affascinò molto:
Una volta, in un seminario del nostro dottorato ci venne a trovare un professore che provò a spiegare cosa fosse e dove si trovasse la nostra coscienza. Non ricordo nulla di interessante in quelle parole, ma una cosa mi affascinò molto:
Nel primo anno di vita, un bambino, con tutto il rispetto, è molto simile agli animali per quanto riguarda la coscienza della propria esistenza nel mondo. Lo si può notare quando portate un bambino di circa 6 mesi di fronte ad uno specchio, noterete che il primo approccio è quello di salutare, afferrare o interagire con l’immagine sua riflessa senza rendersi conto che quell’individuo risponde ai propri movimenti. In seguito ad una esposizione più lunga i bambini mostrano una quasi totale indifferenza a quello strano vetro magico, in pratica capiscono che è un trucco e non tentano più una interazione. L’approccio dei classici animali domestici di frone al ‘vetro magico’ é lo stesso. Chi ha un cane o un gatto avrà sicuramente notato una interazione in una prima fase, poi una totale indifferenza.
Nell’uomo però c’è una rivoluzione sostanziale che, se non fosse blasfemo, potremmo definire miracolosa. Un bambino, tra i sei mesi fino ad un anno di età, proprio nella fase in cui comincia a parlare, camminare e fare alcuni abbozzi che ripeterà continuamente da grande, ha anche la rivoluzione di capire che esiste nel mondo. Portato al ‘vetro magico’ questo bambino ‘cresciuto’, ha una interazione con lo specchio del tutto diversa… comincia a capire che quello che vede è se stesso, a toccarsi nelle zone che ritiene diverse dal solito, es macchie evidenti, sporco, colori che non gradisce. Anche di fronte alla televisione incomincia uno spirito di identificazione ed imitazione spinto proprio dai suoni.
Link video Spirito di Imitazione
Per quanto si sappia, questa è una caratteristica di pochissime specie animali, solo alcune scimmie e l’uomo (vedi post “Allo specchio“). Tuttavia solo la vista e l’udito possono stimolare questo istinto naturale.
Non vorrei entrare nel merito di questa strana e misteriosa maturazione nervosa che è ancora più colossale della nascita stessa del bambino, poiché non sto trattando solo di coscienza, linguaggio, equilibrio, voglia di interazione sociale ed una maturazione anche del carattere che avrà da grande, parlo anche di capacità di capire i suoni e dare a questi un significato. Insomma la rottura definitiva tra il vecchio mondo ancestrale e quello più evoluto e complesso degli esseri umani sapiens sapiens, ricco di emozioni, poesie, pensieri, riflessioni e rancori.
Nel prossimo post vedremo uno di questi piccoli ‘miracoli’ della vita, un argomento di neuroscienze, che è avvenuto in tanti bambini senza che ce ne rendiamo conto.
Conclusione
Qualcuno disse che “non è possibile cogliere un fiore senza turbare una stella” oppure “una farfalla che sbatte le ali in brasile crea una tempesta in un altro continente”. Bhé insomma, questo che abbiamo visto è un piccolo ‘dettaglio storico’ rappresentato dalla farfalla, nel prossimo post vedremo la perturbazione che ne consegue sul campo delle neuroscienze.
Continua: L’Orecchio: Assoluto o Relativo? (Parte II)
Tags:
Neuroscienze,
Percezione,
suono,
udito
15 settembre 2009 - 6:48 pm
Qualcuno disse che non ci può essere giallo dove non ci sia anche il maggiordomo e la servitù. Il nostro poliziesco non ne è immune ovviamente, vediamo il proseguio delle indagini.
Preambolo
Continuiamo il nostro percorso in questo strano giallo, per chi non avesse seguito le puntate precedenti vi prego di visitare i seguenti links 1, 2, 3 e 4. Qui tratteremo un post di ‘transizione’, molto più semplice dei precedenti, ma che ci serve per capire i prossimi capitoli che saranno un po’ più complessi del solito.
Data la lunghezza eccessiva ho deciso di tagliare la storia in più sezioni, in questa vi presento la premessa (l’incontro). Come al solito sono bene accetti commenti di ogni tipo, suggerimenti per gli argomenti da trattare o migliorare.
La quiete dopo la tempesta
 Come abbiamo visto nel precedente post, il nostro ispettore cellulare è stato improvvisamente freddato dallo scagionamento del primo imputato eccellente. La teoria iniziale non faceva una grinza e c’erano anche molti indizi pesanti contro l’accusato. Purtroppo con il tempo e con l’uso di sostanze sempre più specifiche per il blocco del fattore transcrizionale la teoria della difesa ha avuto sempre più supporto. HIF-1 non è sicuramente una proteina da mettere tra i cattivi per l’ischemia cerebrale. A quanto pare c’è stato un grosso fallimento da cui abbiamo imparato a non concludere facilmente le indagini prima di approfondirle con cura.
Come abbiamo visto nel precedente post, il nostro ispettore cellulare è stato improvvisamente freddato dallo scagionamento del primo imputato eccellente. La teoria iniziale non faceva una grinza e c’erano anche molti indizi pesanti contro l’accusato. Purtroppo con il tempo e con l’uso di sostanze sempre più specifiche per il blocco del fattore transcrizionale la teoria della difesa ha avuto sempre più supporto. HIF-1 non è sicuramente una proteina da mettere tra i cattivi per l’ischemia cerebrale. A quanto pare c’è stato un grosso fallimento da cui abbiamo imparato a non concludere facilmente le indagini prima di approfondirle con cura.
Siamo quindi costretti a ritornare sui nostri passi ed ricominciare le indagini dall’inizio con più zelo, cercando di raccogliere informazioni sempre più precise e concordanti su cosa stava accadendo ai neuroni poco prima di morire.
Direi di iniziare l’interrogatorio su tutti i possibili testimoni oculari o detentori di informazioni cruciali.
Dove trovare i colpevoli?
 Pensare all’ipossia come innesco di un meccanismo che porterà a morte inevitabile per i neuroni non è male come teoria, tuttavia ci sono dei ricercatori che la pensano diversamente. Una volta, in un congresso un mio amico mi fece un’obbiezione inappellabile… per quanto possa essere interessante quel meccanismo che innesca la morte delle cellule, non c’è modo di evitarlo, un paziente arriva al pronto soccorso quando manifesta già i sintomi, ovvero quando c’è già stato “l’innesco letale”. Inoltre non è neanche possibile prevedere quando un individuo avrà un ictus, la nostra unica possibilità di aiutarlo risiede proprio in quella finestra di tempo che intercorre tra la comparsa dei sintomi e l’effettiva morte neuronale.
Pensare all’ipossia come innesco di un meccanismo che porterà a morte inevitabile per i neuroni non è male come teoria, tuttavia ci sono dei ricercatori che la pensano diversamente. Una volta, in un congresso un mio amico mi fece un’obbiezione inappellabile… per quanto possa essere interessante quel meccanismo che innesca la morte delle cellule, non c’è modo di evitarlo, un paziente arriva al pronto soccorso quando manifesta già i sintomi, ovvero quando c’è già stato “l’innesco letale”. Inoltre non è neanche possibile prevedere quando un individuo avrà un ictus, la nostra unica possibilità di aiutarlo risiede proprio in quella finestra di tempo che intercorre tra la comparsa dei sintomi e l’effettiva morte neuronale.
Se ci pensate è assurdo che le cellule siano ancora vive quando arrivano nelle nostre mani, e nonostante i nostri mezzi siamo incapaci evitarne la morte. Se vogliamo veramente fare qualcosa il primo punto dell’ordine del giorno deve essere fermare l’arma che ucciderà i neuroni, e solo dopo capire chi ha innescato il meccanismo ed è colpevole (scoprire il mandante).
Nel nostro caso dovremmo soffermarci sul perché le cellule non riescono più a gestire il calcio e muoiono. La soluzione di questo enigma sarà certamente correlata con la possibilità di salvare le cellule, mentre la cattura del mandante che ha scatenato la serie di eventi non servirà ad evitare nessun decesso cellulare al momento. Solo in un passaggio successivo potremmo chiarire anche chi ha causato tale massacro neuronale e perché.
Seguendo questa linea di pensiero in questo post ci concentreremo sui meccanismi di controllo e mantenimento dell’omeostasi ionica, cercando di fare il punto della situazione per poi trarne degli indizi.
La prima domanda è chi mantiene in ordine le concentrazioni ioniche intra ed extra cellulari?
 Se fate questa domanda ad un qualsiasi esperto nel settore vi dirà che ci sono migliaia di proteine che fanno ‘ordine’ tra i diversi ioni, e sono divise in famiglie, superfamiglie e classi. Questi meccanismi possono dividersi grossolanamente in pompe e trasportatori, in base al fatto che usano direttamente, o no, l’ATP.
Se fate questa domanda ad un qualsiasi esperto nel settore vi dirà che ci sono migliaia di proteine che fanno ‘ordine’ tra i diversi ioni, e sono divise in famiglie, superfamiglie e classi. Questi meccanismi possono dividersi grossolanamente in pompe e trasportatori, in base al fatto che usano direttamente, o no, l’ATP.
Tra le pompe più famose ci sono la Na+/K+ ATPasi che estrude sodio e fa entrare potassio, la pompa del calcio di membrana ed endoplasmatica (PMCA e SERCA), etc. Tra i trasportatori passivi i più noti sono lo scambiatore sodio/calcio (NCX), lo scambiatore sodio/calcio potassio (NCKX), lo scambiatore sodio/idrogeno (NHE), ed altri. Si tratta di un gruppo enorme di proteine che mettono continuamente in ordine i vari ioni, riportando continuamente le condizioni cellulari allo stato di riposo dopo le stimolazioni nervose che sono servite per attivare la cellula e per comunicare con il neurone successivo. E’ un po’ come avere delle cameriere e dei maggiordomi in una grande villa che mettono continuamente in ordine le camere ogni qual volta sono utilizzate.
Oggi ci concentreremo proprio su questi personaggi che stavano lì quando dalla calma apparente si è scatenata la disfatta neuronale.
La domanda ora è ‘Perché non hanno funzionato a dovere nel momento del bisogno?’
Vale la pena rispondere con chiarezza.
Prima domanda: Chi era in servizio durante l’ischemia e poco prima?
 A questa domanda c’è una risposta più o meno ovvia. Poco prima dell’ischemia funzionava tutto come al solito, nulla lasciava presagire il disastro, e tutte le proteine risiedevano al loro nomale posto fino a che è mancata l’energia necessaria per il trasporto attivo degli ioni. Quando è iniziata a mancare l’energia l’efficienza del trasporto ionico si è ridotta mentre il disordine ionico dei neuroni aumentava. Poco dopo questa fase gli unici che potevano fare qualcosa erano gli scambiatori ionici che non necessitano di ATP, ma il destino della cellula era oramai segnato; senza ATP non si può andare avanti per molto. Poi d’improvviso è arrivata nuova energia e tutto è cominciato a rifunzionare alla normalità fino al danno irreparabile.
A questa domanda c’è una risposta più o meno ovvia. Poco prima dell’ischemia funzionava tutto come al solito, nulla lasciava presagire il disastro, e tutte le proteine risiedevano al loro nomale posto fino a che è mancata l’energia necessaria per il trasporto attivo degli ioni. Quando è iniziata a mancare l’energia l’efficienza del trasporto ionico si è ridotta mentre il disordine ionico dei neuroni aumentava. Poco dopo questa fase gli unici che potevano fare qualcosa erano gli scambiatori ionici che non necessitano di ATP, ma il destino della cellula era oramai segnato; senza ATP non si può andare avanti per molto. Poi d’improvviso è arrivata nuova energia e tutto è cominciato a rifunzionare alla normalità fino al danno irreparabile.
In questa indagine tutto è sottoposto alla lente di ingrandimento, e nulla è dato per scontato, quindi le dichiarazioni vanno convalidate e come al solito andiamo in laboratorio con una ricostruzione del delitto per cercare di confutare tali ipotesi. In questo caso potremmo ricorrere ad esperimenti in vitro fatto con delle cellule neuronali sottoposte a dei modelli di deprivazione di ossigeno e glucosio, seguito dalla riossigenazione. Si tratta ovviamente di un modello molto semplice, ma che ci evita di sacrificare animali, e consente un maggiore grado di controllo delle condizioni sperimentali. Effettivamente quando manca ATP funzionano solo i trasportatori passivi, e la loro capacità di controllare l’omeostasi ionica è discreta considerando le condizioni di emergenza. Inoltre nulla sembra cambiare nell’assetto dei vari trasportatori attivi e passivi, dopotutto non c’è abbastanza energia e tempo per cambiare le cose con una sintesi/degradazione proteica. I sistemi sono ri-settati all’essenziale ed a tutto quello che può aiutare a tamponare i danni o trarre energia.
Se tutte queste considerazioni sono vere dobbiamo escludere chi non poteva funzionare al momento dell’ictus e considerare invece solo chi poteva ed ha fatto qualcosa…
La prossima domanda è quindi: Chi funzionava al momento e cosa stava facendo?
NCX: il maggiordomo
 A questo punto del racconto io mi recherei sul luogo del delitto alla ricerca di qualche indizio e soprattutto di qualche risposta.
A questo punto del racconto io mi recherei sul luogo del delitto alla ricerca di qualche indizio e soprattutto di qualche risposta.
Escludendo le pompe che fanno uso di ATP, tra i pochi trasportatori passivi che rimangono e che possono dare un consistente aiuto c’è lo scambiatore sodio/calcio (NCX).
Io immaginerei un ipotetico interrogatorio a questa proteina, magari in una stanza buia e fumosa.
Lei chi è?
NCX è una proteina di membrana fatta da circa 1.000 aminoacidi che è presente in tutte le cellule dei mammiferi e che fisiologicamente espelle il calcio citosolico in cambio dell’ingresso di sodio.
Da quanto tempo è in servizio?
In pratica quasi tutti gli organismi possiedono uno scambiatore del calcio, e sono tutti filogeneticamente collegati da un gene ancestrale che si è mantenuto molto conservato sia nella sua struttura che nella sua funzione. In altre parole è una proteina presente negli archeobatteri, funghi, piante, fino a tutte le cellule eucariotiche di mammifero, quindi una proteina davvero molto conservata…
Davvero notevole, ma mi dica… qual è la Sua funzione nel cervello e con chi collabora solitamente?
Sappiamo bene che, quando i neuroni si trovano a riposo, le concentrazioni ioniche sono mantenute soprattutto da pompe che fanno uso di energia sottoforma di ATP. In queste condizioni un trasportatore passivo come NCX funziona relativamente poco, poiché ha una bassa affinità per il calcio e per il sodio. Tuttavia le condizioni cambiano drasticamente dopo l’eccitazione neuronale, ovvero quando le concentrazioni di calcio citosolico aumentano improvvisamente a dismisura, le pompe si saturano e non riescono a riportare la cellula alle condizioni ioniche di riposo. In queste circostanze entrano in funzione NCX e proteine simili che hanno una bassa affinità per gli ioni, ma possono trasportarne enormi quantità in pochi istanti.
In pratica NCX espelle uno ione calcio mediante scambio con tre ioni sodio che entrano nella cellula, (proprio da questo deriva il suo nome).
Quindi Lei è specializzato nelle condizioni di ‘emergenza’, ovvero quando le pompe non riescono a mettere ordine… cosa mi sa dire riguardo all’ipossia? cosa stava facendo in quei momenti?
La risposta sarebbe ovvia. NCX stava espellendo il calcio dalle cellule sfruttando l’unica fonte di energia disponibile al momento ovvero il gradiente del sodio.
Perché?
Come già abbiamo accennato nei post precedenti il calcio è un ottimo segnale citosolico, ma quando supera certi livelli può causare disastri, quindi l’azione di NCX è in linea con le aspettative.
Ha visto qualcosa di strano in quei ‘momenti’? Può dirmi qualcosa a tal riguardo?
Quello che NCX ha visto è che la pompa del calcio in una prima fase era efficiente e riusciva a mantenere le concentrazioni di calcio citosolico a livelli estremamente bassi, ma riusciva ad espellere solo pochi ioni alla volta. Ad un certo punti si stava accumulando il calcio, la pompa è andata subito in saturazione e non riusciva ad espellere abbastanza ioni… poi è cominciata a mancare l’energia e l’efficienza di estrusione si è ridotta sempre di più. Si stava accumulando calcio citosolico oltre i normali livelli e tempi sopportabili per il neurone, per cui NCX è entrato in funzione ed ha espulso fino ad oltre 10.000 volte più ioni calcio nell’unità di tempo rispetto alla pompa in presenza di ATP, un fenomeno insomma.
…e cosa sa dirmi su cosa sia successo negli ultimi ‘istanti di agonia’? Alla fine è riuscito a tenere sotto controllo il calcio?
Effettivamente ci è riuscito parzialmente, poiché durante l’ipossia le concentrazioni di calcio si sono mantenute più o meno stabili per quanto fosse possibile farlo senza utilizzare ATP, e soprattutto fatto in così poco tempo… successivamente al ritorno dell’ossigeno tutto è tornato ad una apparente normalità; il livello di calcio citosolico è tornato nel suo normale range di concentrazioni citosoliche, quindi tutto ok, passata la tempesta è stato ovvio pensare che il peggio era oramai passato. Tuttavia, dopo un po’ qualcosa è cominciato ad andare sempre peggio, ci sono stati degli sbalzi di calcio improvvisi sempre più gravi, e la cellula si è resa conto ad un certo punto che non era possibile recuperare la situazione e si è arrivati all’inevitabile. Quei momenti sono stati talmente turbolenti che non è stato possibile capire cosa stesse succedendo.
Uhmm.. Per ora l’interrogatorio è finito, e diciamo che siamo soddisfatti delle informazioni raccolte…
Riepilogando, stiamo parlando di una proteina molto conservata, al che se ne potrebbe dedurre che il suo ruolo sia fondamentale per la sopravvivenza; insomma sembra interessante, ma purtroppo non sembra essersi aperta alcuna strada per noi. Praticamente non ci ha detto nulla.
C’è un’altra cosa…
 Ritornando in ufficio c’è una cosa che non quadra… questo trasportatore passivo dove ha trovato tutta questa energia per espellere una quantità di calcio ionico così colossale? Una pompa del calcio consuma circa 1 ATP per 1 ione calcio, possibile che NCX abbia lavorato gratis contro ogni legge della termodinamica? In altre parole quanto è costato alla cellula questo ‘servizio’?
Ritornando in ufficio c’è una cosa che non quadra… questo trasportatore passivo dove ha trovato tutta questa energia per espellere una quantità di calcio ionico così colossale? Una pompa del calcio consuma circa 1 ATP per 1 ione calcio, possibile che NCX abbia lavorato gratis contro ogni legge della termodinamica? In altre parole quanto è costato alla cellula questo ‘servizio’?
Ovviamente NCX ha sfruttato il gradiente di sodio tra l’ambiente extracellulare e quello intracellulare, ed in questo modo ha potuto espellere il pericoloso calcio dalla cellula scambiandolo con l’innocuo sodio…
Ma cosa succede a tutto quel sodio che NCX ha fatto entrare nella cellula e che non può essere espulso dalla pompa sodio/potassio?
Se sono stati necessari 3 ioni sodio per espellere ciascuno ione calcio, facendo dei semplici calcoli, a naso direi che nella cellula si sarà accumulato tanto di quel sodio da innescare altri meccanismi, ad esempio alterare il potenziale di membrana.
Se questo fosse vero la depolarizzazione causerebbe l’attivazione della cellula, con rilascio di neurotrasmettitori e conseguenze da definire. Sicuramente potremmo parzialmente spiegare l’ipereccitabilità, il calcio che rientra nuovamente nelle cellule e le onde di eccitazione neuronale che si espandono dal core ischemico.
La situazione non è molto chiara, e tornando alla centrale io darei un’occhiata allo schedario delle fedine penali, www.pubmed.gov , in cerca di cosa si sappia in giro di questo ‘maggiordomo’.
Da una rapida occhiata possiamo vedere che è correlato con numerosi meccanismi, NCX si lega con pompe e trasportatori ed esalta alcune caratteristiche dei canali ionici. Però qualcosa è più importante di tutto, è stato visto correlato con l’ischemia cardiaca; leggendo bene possiamo vedere che l’uso di farmaci che inibiscono l’attività di NCX diminuisce l’estensione della morte del miocardio in seguito all’infarto.
L’ipotesi è che la cellula si sia talmente indebitata di energia durante l’ipossia che al ripristino delle condizioni ottimali non ce n’è abbastanza per ristabilire il gradiente di sodio e reimpostare le condizioni di riposo. Un giro vizioso che può portare solo al disastro, come aver chiesto dei soldi ad uno strozzino.
Una crepa sul vetro cristallino di NCX: Calunnie?
 Uhmm… Forse calunnie, leggendo bene qua e là i farmaci utilizzati per inibire NCX non sono molto specifici, agiscono anche su altri canali; magari NCX è stato coinvolto in qualcosa di più grande ed è stato usato come capro espiatorio. Improbabile che una proteina così letale possa essere conservata per milioni di anni durante l’evoluzione se non migliori sensibilmente il fitness dell’organismo.
Uhmm… Forse calunnie, leggendo bene qua e là i farmaci utilizzati per inibire NCX non sono molto specifici, agiscono anche su altri canali; magari NCX è stato coinvolto in qualcosa di più grande ed è stato usato come capro espiatorio. Improbabile che una proteina così letale possa essere conservata per milioni di anni durante l’evoluzione se non migliori sensibilmente il fitness dell’organismo.
Per dare una prova decisiva possiamo utilizzare, come tutti voi sapete, la prova del nove… un topo transgenico knock-out per NCX e verificare cosa succede.
In questo modello, infatti, se non ci sono variazioni dell’espressione degli altri canali/trasportatori c’è la massima specificità dell’inibizione di questa proteina… e quindi potremmo ufficialmente capire se si tratta di una proteina così cattiva.
Andiamo in lab e chiediamo di sviluppare il topo knock-out. Passano anni e qui il primo intoppo, il topo muore durante l’embriogenesi per mancato battito cardiaco. Si tratta di una scoperta senz’altro interessante ma sono anche anni di lavoro buttati al vento senza una risposta. Rifare tutto il lavoro daccapo e passare altri anni a sviluppare un altro topo? Uhmm…
Qualcuno ha suggerito una soluzione da fantascienza che potrebbe risolvere la situazione, ovvero utilizzare un virus in grado di infettare i cardiomiociti dell’embrione knock-out ed inserire in queste cellule il cDNA di NCX in maniera transiente… insomma abbastanza per attivare il battito cardiaco, poi successivamente l’espressione calerà di nuovo quando il topo non sarà più infettato.
Lo sviluppo della metodologia ha impiegato un paio di anni con risultati sconfortanti, il risultato è stato che l’espressione transiente di NCX nei cardiomiociti non serve a nulla. La mancanza di NCX è fondamentale per la vita dell’animale, probabilmente è necessaria l’espressione anche in altri tessuti (es le cellule pacemaker del cuore, di tipo nervoso), oppure è necessaria una espressione ancora più precoce della proteina, magari quando non si è ancora formato il primo abbozzo di cuore.
Disperati, si è ricorsi a strategie farmacologiche sempre più complesse, ma i risultati sono stati discordanti e dubbi; mancano sempre di selettività e specificità.
Il volo Pindarico
 Le cose, da allora, non sono andate molto avanti, le pubblicazioni si sono rincorse tra di loro in un misto di ripetizioni e staccate con farmaci sempre più specifici e potenti. A questi articoli facevano seguito altri che descrivevano dell’effetto di queste sostanze sulla circolazione, ipertensione, inibizione di canali del calcio, respirazione mitocondriale, disaccoppiamenti mitocondriali etc etc.
Le cose, da allora, non sono andate molto avanti, le pubblicazioni si sono rincorse tra di loro in un misto di ripetizioni e staccate con farmaci sempre più specifici e potenti. A questi articoli facevano seguito altri che descrivevano dell’effetto di queste sostanze sulla circolazione, ipertensione, inibizione di canali del calcio, respirazione mitocondriale, disaccoppiamenti mitocondriali etc etc.
Tuttavia per fare una buona ricerca c’è bisogno di determinazione, genialità ed anche un po’ di fortuna, tutte caratteristiche ben rappresentate nel padre di NCX, Kenneth Philipson, quando a sorpresa decide di intraprendere un nuovo e più Pindarico volo, investendo diversi anni nella generazione di un nuovo topo knock-out per NCX, ma questa volta condizionato (tecnica Cre/LoxP). Si tratta di una tecnica citata già più volte e che funziona bene, in poche parole il topo nascerà con una normale espressione di NCX in tutto l’organismo, poi ad un certo punto l’enzima Cre, come una bomba ad orologeria, eliminerà il gene di NCX solo nei cardiomiociti e darà un topo adulto privato improvvisamente di NCX.
Per i più curiosi il meccanismo sfrutta il cambio della catena pesante della miosina dei cardiomiociti subito dopo la pubertà. In questi topi transgenici insieme al promotore della catena pesante della miosina c’è una ricombinasi che andrà a distruggere il gene ncx causando un knock-out tessuto e tempo specifico.
Non vi dico del rischio che il topo muoia proprio quando la proteina verrà a mancare e vanificare ancora una volta il lavoro di anni di sacrifici.
Passarono degli anni e finalmente eravamo arrivati vicino al fatidico momento, la nascita del primo topo transgenico. Da quel momento devo dire che c’è stata una lunga agonia misto di silenzio ed impazienza aspettando quel fatidico momento in cui un topolino che ha in sé tutte le promesse di questo mondo deve passare per quell’unico istante in cui diventerà knock-out e dimostrerà al mondo se è possibile vivere senza una specifica proteina cardiaca.
Purtroppo capita anche di affezionarsi ad un animale che vivendo darà inconsapevolmente un enorme contributo alla ricerca. I giorni passavano ed il topo alla sua maturità sessuale era ancora vivo e vitale, le analisi non hanno dimostrato anomalie cardiache e tutto procede come nulla fosse. Così è stato anche per le nidiate successive e la biochimica ha dato l’ok, questi topi adulti hanno un knock-out cardiaco. Si decide di fare l’ischemia cardiaca e il risultato è stato sconcertante. La mancanza di NCX nel cuore provoca un’entità del danno al miocardio pari al 50% circa rispetto ai topi wild-type.
NCX è sicuramente colpevole… un fulmine nel cielo sereno, una pubblicazione che cambierà definitivamente la storia di questo trasportatore.
Su questo modello animale sono stati poi tentati tantissimi esperimenti, soprattutto con quei farmaci così potenti e specifici per NCX; i risultati hanno dimostrato in maniera incontrovertibile che gli effetti ottenuti erano dovuti all’azione su altre proteine, è stato solo un caso che l’uso di queste sostanze dimostrano gli stessi effetti del knock-out per NCX.
Quello che succede al cuore succede anche al cervello?
 Analisi su analisi in diverse salse hanno dimostrato che questa proteina è fondamentale per alcuni meccanismi cardiaci e non solo, ma sicuramente esalta anche il danno ischemico al miocardio. E’ una circostanza difficile da spiegare in termini evoluzionistici e non è possibile capirne a fondo il meccanismo esatto.
Analisi su analisi in diverse salse hanno dimostrato che questa proteina è fondamentale per alcuni meccanismi cardiaci e non solo, ma sicuramente esalta anche il danno ischemico al miocardio. E’ una circostanza difficile da spiegare in termini evoluzionistici e non è possibile capirne a fondo il meccanismo esatto.
Qualcuno fa notare che sia l’eccesso di ioni sodio durante il lavoro di NCX a causare danno per osmosi; tre ioni sodio per miliardi di ioni calcio determinerebbero uno squilibrio osmotico che liserebbe la membrana cellulare formando prima delle bolle sulla superficie e poi la rottura fino alla morte.
Ovviamente la trasposizione al cervello è immediata, se fa male al cuore potrebbe far male anche ai neuroni?
La situazione non è semplice, abbiamo diverse opzioni per verificare questa ipotesi e nessuna di queste è facile.
L’esperimento più determinante potrebbe essere, come al solito, la generazione di un knock-out per NCX nel cervello, però si corre nuovamente il rischio di un nuovo tonfo in acqua, poiché potrebbe mancare l’embriogenesi del SNC e quindi aborto precoce. Se questo dovesse avvenire avremmo speso tempo e denaro inutilmente, per non parlare delle sofferenze indotte sugli animali.
D’altra parte fare dei knock-out tessuto specifico, soprattutto per il cervello, è difficile, lungo e dispendioso; per ora accantoniamo l’idea fino a che non sia strettamente necessario.
Potremmo ricorrere a dei saggi alternativi per chiarire la situazione, e magari li vediamo nella prossima puntata.
Conclusione
Si potrebbe definire serendipità: cercare un ago in un pagliaio e trovarci la figlia del contadino (Julius H. Comroe).
Stavamo brancolando nel buio in cerca di prove e ci siamo scontrati con un pregiudicato per il danno cardiaco ischemico camuffato da semplice faccendiere delle pulizie da calcio.
Nelle prossime puntate analizzeremo meglio il personaggio, il suo ruolo nel cuore e poi nel cervello, cercheremo nuove prove a cui ovviamente farà seguito il ‘processo’ per concorso in neuro-omicidio.
Alla prossima
Tags:
Animali transgenici,
Calcio,
Cre/LoxP,
Ictus,
Ischemia Cerebrale,
NCX,
Neuroni,
Neuroscienze,
PMCA,
pompe,
Potenziali d'azione,
sodio
4 settembre 2009 - 1:00 pm
Preambolo
Finalmente le tanto meritate vacanze sono iniziate anche per me, e colgo l’occasione per finire finalmente questo post in elaborazione oramai da tanto tempo, come al solito cortesemente corretto da Valentina. Qui descriverò un argomento abbastanza lungo, difficile e ‘spigoloso’ per i profani ed esperti, anche se penso che tutti lo conoscono almeno un po’ per sentito dire.
Parleremo dell’olfatto, un argomento non facile da raccontare in un post divulgativo senza ‘smussare’ qua e là il mosaico di teorie e di esperimenti che sono stati fatti fino ad ora.
L’Olfatto: Alcuni dettagli ovvi

Come al solito mi servirò inizialmente di qualche punto di riferimento per metterci d’accordo ed inizierò dagli odori che sono causati, come tutti voi sapete, da molecole sprigionate dalle sostanze e disperse nell’aria che respiriamo, dove raggiungono il nostro naso. L’altro punto di riferimento utile si trova subito dietro il naso e sopra la volta della nostra cavità buccale dove passano tutte le molecole che noi respiriamo. E’ una cavità rivestita da una mucosa di circa 10 cm2 che attraverso la solubilizzazione, la dispersione, l’adsorbimento e quant altro, ferma una porzione delle molecole che si trovano nello stadio aeriforme nella cavità nasale e le disperde nel fluido mucoso. Qui le molecole possono incontrare una o più cellule recettoriali olfattive (ORC), in un uomo adulto se ne contano circa 10-20 milioni, ricche di propagini dette ciglia e di proteine dette recettori olfattivi (da qui chiamate Odorant Receptor OR).
I OR hanno il compito ‘legare’ in maniera più o meno specifica alcuni gruppi funzionali di queste molecole presenti nel fluido mucoso, ed innescare una cascata di eventi che si amplifica fino ad attivare la cellula recettrice e scatenare la propagazione del ‘segnale odoroso’. Il cervello, a sua volta, provvederà ad interpretare il segnale in diversi stadi con una serie di meccanismi molto complessi. Per dettagli ed approfondimenti potete guardare questo link.
Fin qui il tutto è quasi scontato ed ovvio, però dalla teoria alla pratica c’è una notevole differenza, vi dico subito che ancora oggi ci sono vari lati oscuri che cercherò di descrivervi in parole semplici, portandovi ai problemi scientifici in maniera intuitiva per quanto sia nelle mie capacità.
Quanti odori esistono?
 Come penso molti di voi possono intuire, esiste un numero inimmaginabile di odori, e la maggioranza di questi è causato da un mix di ‘odori primari’ in diversi rapporti, ma a complicare la situazione c’è che una molecola odorosa può agire su decine di recettori diversi con diversa specificità. I OR dei composti aminici, ad esempio, riconoscono solamente l’azoto primario, secondario o terziario con diversa efficacia, per cui gli odori di questi composti non possono essere discriminati con facilità e sembrano tutti molto simili. C’è da dire però che nonostante questa piccola sovrapposizione di segnali si possono discriminare con elevata sensibilità circa 10.000 odori diversi.
Come penso molti di voi possono intuire, esiste un numero inimmaginabile di odori, e la maggioranza di questi è causato da un mix di ‘odori primari’ in diversi rapporti, ma a complicare la situazione c’è che una molecola odorosa può agire su decine di recettori diversi con diversa specificità. I OR dei composti aminici, ad esempio, riconoscono solamente l’azoto primario, secondario o terziario con diversa efficacia, per cui gli odori di questi composti non possono essere discriminati con facilità e sembrano tutti molto simili. C’è da dire però che nonostante questa piccola sovrapposizione di segnali si possono discriminare con elevata sensibilità circa 10.000 odori diversi.
Ciò premesso, consentitemi da questo punto in poi di semplificare e di concentrarci esclusivamente sugli ‘odori primari‘ che sono riconosciuti quasi esclusivamente da un solo tipo di OR e che danno una sensazione odorosa molto precisa.
Cosa c’è di difficile?
 Nel mondo animale, il non saper distinguere l’odore di una preda dall’odore del proprio predatore fa la differenza tra fare una buona colazione ed essere la colazione di qualcun altro. Anche riconoscere l’odore delle cose che si possono mangiare da quelle che è meglio evitare è fondamentale per la sopravvivenza, quindi i mammiferi durante l’evoluzione hanno accumulato più di 1.000 geni che sovraintendono il più misterioso dei 5 sensi. In pratica più del 3% del genoma dei mammiferi è deputato alla percezione degli odori, e dato che questi recettori possono riconoscere anche più tipologie di molecole con diversa affinità determinando una combinazione di interazioni intermedie, questo amplifica esponenzialmente il ventaglio di sensazioni che possono dare le molecole presenti nell’aria.
Nel mondo animale, il non saper distinguere l’odore di una preda dall’odore del proprio predatore fa la differenza tra fare una buona colazione ed essere la colazione di qualcun altro. Anche riconoscere l’odore delle cose che si possono mangiare da quelle che è meglio evitare è fondamentale per la sopravvivenza, quindi i mammiferi durante l’evoluzione hanno accumulato più di 1.000 geni che sovraintendono il più misterioso dei 5 sensi. In pratica più del 3% del genoma dei mammiferi è deputato alla percezione degli odori, e dato che questi recettori possono riconoscere anche più tipologie di molecole con diversa affinità determinando una combinazione di interazioni intermedie, questo amplifica esponenzialmente il ventaglio di sensazioni che possono dare le molecole presenti nell’aria.
La chiave di volta per ottenere un buon olfatto è sicuramente il collegamento tra le cellule recettoriali odorose (ORC) ed il cervello, ovvero quel meccanismo che consente di poter capire quale specifico OR è stato attivato. Si tratta di un meccanismo per nulla banale, e come punto di riferimento, possiamo considerare l’esempio del gusto, dove i recettori cellulari sono generati in base alla localizzazione sulla superficie della lingua. Nel cervello si forma una sorta di correlazione tra aree specifiche corticali ed aree della lingua denominata ‘mappa sensoriale‘. A questo consegue che è possibile percepire il salato, l’amaro, il dolce ed l’acido in base a quale zona della lingua è stata stimolata maggiormente.
Possiamo trasportare questa ipotesi nell’olfatto, ed immaginare che l’area in cui si trovano le ORC determina anche il set di recettori da esprimere. Ipotizziamo che ciascuna cellula recettrice dell’olfatto possa esprimere un centinaio o più OR localizzati in diverse aree della mucosa nasale. Ogni area in cui vi sono le ORC potrebbe essere correlata ad una zona del cervello a cui corrisponderebbero determinate sensazioni odorose.
Quanti recettori per cellula?
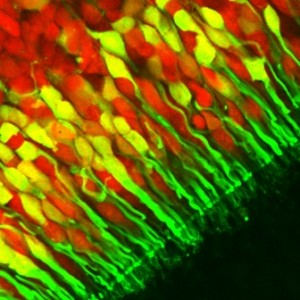 Sapere quanti tipi di recettori sono presenti per cellula è fondamentale per capire quante aree ci dovremmo aspettare di trovare e per verificarlo potremmo ricorrere a tecniche avanzatissime di in situ hybridization, e di retrotrascrizione ed amplificazione per PCR su singola cellula. Ovviamente non sono tecniche facili e nemmeno tanto diffuse, ma gli esperti sono riusciti a dimostrare con questi esperimenti che ogni ORC esprime solamente un singolo OR.
Sapere quanti tipi di recettori sono presenti per cellula è fondamentale per capire quante aree ci dovremmo aspettare di trovare e per verificarlo potremmo ricorrere a tecniche avanzatissime di in situ hybridization, e di retrotrascrizione ed amplificazione per PCR su singola cellula. Ovviamente non sono tecniche facili e nemmeno tanto diffuse, ma gli esperti sono riusciti a dimostrare con questi esperimenti che ogni ORC esprime solamente un singolo OR.
Una cosa simile avviene durante il differenziamento cellulare, dove la combinazione di alcuni fattori trascrizionali decide l’attivazione di meccanismi sempre più specializzati che ‘bloccano’ l’espressione specifica di determinati geni. Questo meccanismo, definito ‘deterministico’, è quello maggiormente utilizzato durante la specializzazione cellulare nel nostro organismo. Potremmo ipotizzare un meccanismo simile per le ORC in cui dalla combinazione di alcuni fattori trascrizionali si determina l’attivazione dell’espressione di un singolo OR.
Verifica della Teoria Deterministica
 Secondo questa teoria la combinazione casuale o pseudocasuale di fattori trascrizionali per esprimere un singolo OR non può essere disturbata dalla coespressione di un OR transgenico scelto da noi. Per verificare questa teoria ‘deterministica‘, quindi, possiamo generare un topo transgenico in cui alcune ORC esprimono in maniera forzata un determinato OR scelto da noi e valutare se c’è coespressione.
Secondo questa teoria la combinazione casuale o pseudocasuale di fattori trascrizionali per esprimere un singolo OR non può essere disturbata dalla coespressione di un OR transgenico scelto da noi. Per verificare questa teoria ‘deterministica‘, quindi, possiamo generare un topo transgenico in cui alcune ORC esprimono in maniera forzata un determinato OR scelto da noi e valutare se c’è coespressione.
Il risultato è interessante, non c’è alcuna sovrapposizione di segnali tra il OR espresso forzatamente da noi ed un qualsiasi altro recettore olfattivo endogeno. In pratica l’espressione forzata del nostro OR transgenico non permette di esprimere un qualsiasi altro OR endogeno, e se si tenta di generare un topo transgenico con una espressione forzata di due OR diversi nella stessa ORC, queste muoiono per apoptosi. Questi dati non sono di certo a supporto per la teoria deterministica, evidentemente la scelta del recettore olfattivo è un meccanismo molto più complesso. Ad aggiungere benzina sul fuoco c’è un’altra prova interessante, ovvero le ORC possono cambiare più volte il recettore odoroso espresso prima di prendere contatto con il cervello (maturazione cellulare). La teoria deterministica è molto fallace con questi risultati, poco probabile che sia quella buona.
La Teoria Stocastica
 Un’altra teoria che si è fatta strada nel tempo è basata sulla casualità della scelta del OR espresso (teoria stocastica). Per capirci immaginiamo un fattore trascrizionale unico che sia in grado di legare il promotore di tutti i geni dei OR e di consentirne l’espressione; tecnicamente può legarsi ad un solo OR e quindi può esprimere solo quello. Se il fattore trascrizionale non è molto efficiente nell’esprimere un OR si potrebbe spiegare come mai una ORC cambia OR nei primi stadi pur seguendo sempre la regola di esprimere un solo OR alla volta. In seguito, quando un OR espresso dà un forte segnale di funzione (feed-back) il fattore trascrizionale si fissa sul gene che sta trascrivendo fino allo stadio di maturazione.
Un’altra teoria che si è fatta strada nel tempo è basata sulla casualità della scelta del OR espresso (teoria stocastica). Per capirci immaginiamo un fattore trascrizionale unico che sia in grado di legare il promotore di tutti i geni dei OR e di consentirne l’espressione; tecnicamente può legarsi ad un solo OR e quindi può esprimere solo quello. Se il fattore trascrizionale non è molto efficiente nell’esprimere un OR si potrebbe spiegare come mai una ORC cambia OR nei primi stadi pur seguendo sempre la regola di esprimere un solo OR alla volta. In seguito, quando un OR espresso dà un forte segnale di funzione (feed-back) il fattore trascrizionale si fissa sul gene che sta trascrivendo fino allo stadio di maturazione.
E’ una teoria che funziona e che spiega anche i risultati dei topi transgenici; il OR transgenico blocca l’espressione di altri recettori attraverso un feed-back abberrante. Però, come al solito la teoria va convalidata, e c’è l’imbarazzo della scelta sul come; una prova schiacciante potrebbe derivare dall’eliminazione del feed-back recettoriale. In particolare potremmo alterare un gene OR in modo tale da codificare un recettore non funzionante pur conservandone la struttura genica.
I risultati dimostrano che il topo transgenico non esprime mai un OR non funzionante, probabilmente proprio per la mancanza del feed-back del recettore maturo necessario per fissare la scelta. Per esserne sicuri, è stata effettuata una controprova per dimostrare che le ORC tentano di esprimere il OR non funzionante, e che solo successivamente cambiano la scelta su un altro gene OR che funzioni. Per dimostrarlo si è generato un topo transgenico in cui l’espressione anche transiente del recettore non funzionante provoca un danno irreversibile al DNA della stessa cellula (tecnica Cre/LoxP), ed i risultati hanno mostrato che le ORC scelgono il OR transgenico e non funzionante con la stessa probabilità di tutti gli altri OR, sebbene questa scelta sia cambiata prima della maturazione cellulare per mancanza di feed-back. Un’altra prova conclusiva e schiacciante è che la generazione di un topo trangenico che esprime un OR esogeno e non funzionante non disturba l’espressione dei OR endogeni. Questo dimostra che è la funzione di un OR a bloccare l’espressione dei OR endogeni, non la struttura e l’espressione dei geni OR endogeni.
La spina nel fianco per la Teoria Stocastica
La teoria stocastica fino ad ora ha sempre soddisfatto tutti i dati sperimentali ottenuti, ma ciononostante ha un punto debolissimo. Questa teoria è retta dalla possibilità di poter innescare la trascrizione di qualsiasi dei 1.000 geni dei OR da uno o pochi fattori trascrizionali, e questo sarebbe possibile solo nel caso in cui i geni OR presentino una sequenza consenso che li identifichi nel genoma. Ebbene, nonostante molti ricercatori si siano impegnati nell’analisi di omologie di sequenze tra i geni degli OR, i risultati ottenuti sono stati alquanto deludenti; con i dati attuali si potrebbero raggruppare non più di una decina di geni degli OR per volta tirando in ballo meccanismi di riconoscimento improbabili.
E se fossero dei Linfociti?
 Se ci pensate, esiste un meccanismo di scelta molto simile che avviene nei linfociti. Queste cellule, però, non decidono l’anticorpo da esprimere in base a fattori trascrizionali, ma attraverso una ricombinazione genetica guidata. Si tratta di una piccola deviazione della teoria stocastica ed è anche risolutiva.
Se ci pensate, esiste un meccanismo di scelta molto simile che avviene nei linfociti. Queste cellule, però, non decidono l’anticorpo da esprimere in base a fattori trascrizionali, ma attraverso una ricombinazione genetica guidata. Si tratta di una piccola deviazione della teoria stocastica ed è anche risolutiva.
Come al solito dobbiamo convalidare tale teoria… ed escludiamo subito la proposta di sequenziare l’intero genoma di singole cellule, sarebbe difficilissimo e ci vorrebbero tanti anni! Possiamo ricorrere ad un trucco chiamato FISH (fluorescent in situ hybridization), ovvero rendere fluorescente una sonda ad RNA verso un introne di un OR. In questa tecnica non è possibile valutare una ricombinazione genica in maniera diretta, tuttavia è possibile localizzare dove inizia la trascrizione di un determinato gene all’interno del nucleo e valutare se, in seguito ad una ricombinazione, cambia posizione. I risultati, alquanto discutibili, dimostrano che il sito di inizio della trascrizione avviene sempre nello stesso punto del nucleo e del genoma, quindi niente ricombinazione, ma c’è una novità inaspettata; la trascrizione di un OR proviene solamente da un singolo allele.
Sbrogliare la matassa di informazioni non è mai facile

Spesso nella ricerca è importante definire delle ipotesi e porsi le domande giuste, ovvero cercare di identificare la cosa più importante di un meccanismo e trarne quante più informazioni possibili per verifica e deduzione. Nel nostro caso la codifica di un singolo allele, cioè solo 1 delle due copie genetiche di un OR, è ancora in linea con la teoria della ricombinazione genetica, poiché se è vero che la scelta è casuale, questa non può avvenire in entrambi gli alleli fratelli allo stesso modo. Inoltre se l’inizio della trascrizione avviene sempre nello stesso punto, questo non esclude che tutto il resto del DNA (es il promotore) possa essere stato ricombinato.
L’ipotesi che ne scaturisce è questa: La cellula sceglie un OR da esprimere in modo casuale, in mancanza di un feed-back funzionale cambia scelta, in seguito, alla maturazione cellulare la scelta diventa definitiva mediante ricombinazione di un promotore o di segnali a monte dell’OR scelto. Questa ipotesi potrebbe spiegare come mai l’espressione forzata di un recettore esogeno possa bloccare l’espressione dei recettori endogeni; probabilmente la proteina inserita provoca un feed-back che innesca una ricombinazione abberrante del DNA nei OR endogeni. La teoria per ora tiene ma con le parole non si dimostra mai niente, è fondamentale convalidare la teoria e questa volta non è per nulla facile evidenziare tante piccole ricombinazioni in singole cellule di 20 micron di diametro circa.
Come dimostrare una ricombinazione che si possa trovare in un qualsiasi punto del genoma? In questo caso il test di validazione è non semplicissimo. L’idea è che la ricombinazione del DNA in una ORC provocherebbe il blocco dell’espressione di un determinato OR in tutte le condizioni, come avviene per un linfocita. Se da questo DNA ricombinato si clonasse il topo stesso da cui deriva… l’animale che risulterebbe dovrebbe esprimere solo un OR, poiché il suo DNA è già ricombinato in maniera irreversibile. I risultati sono stati scioccanti, il topo che ne risulta è perfettamente sano e ha tutto l’apparato olfattivo intatto e funzionante come in un qualsiasi wild-type. Nonostante l’analisi di circa 100 kb a monte ed a valle del gene incriminato non c’è stata nessuna modifica genetica o epigenetica.
Basta un soffio ed il castello crolla
 Certamente la ricombinazione è da scartare ma fino ad ora niente ha smentito la casualità della scelta del OR. Tuttavia un gruppo di ricerca ha recentemente messo in discussione anche questa ipotesi granitica.
Certamente la ricombinazione è da scartare ma fino ad ora niente ha smentito la casualità della scelta del OR. Tuttavia un gruppo di ricerca ha recentemente messo in discussione anche questa ipotesi granitica.
L’idea di fondo è la seguente… Se è vero che la cellula sceglie casualmente un gene su circa 1.000, e di questo solo 1 allele, ne consegue che la scelta è di 1 sui 2.000 alleli disponibili. Se riuscissimo ad inattivare solo 1 di questi 2.000 alleli, ad esempio in modo tale che esprima irreversibilmente EGFP (tecnica Cre/loxP), potremmo rendere fluorescenti le cellule che hanno ‘cambiato idea’ per mancanza di feed-back e verificare quale altro gene sceglie dei 1.999 rimanenti.
Il risultato, ancora una volta, è assolutamente strabiliante. Il topo transgenico presenta una coespressione di EGFP solo con il suo allele funzionante corrispondente. In poche parole l’espressione di EGFP, come previsto, non dà feed-back positivo per cui la cellula cambia OR da esprimere; ma la seconda scelta non è per niente casuale, qualcosa guida la scelta sull’allele di ‘backup’ presente sul cromosoma fratello e non sugli altri 1.998 alleli rimanenti. Come potrebbe la cellula riconoscere in modo specifico l’altro allele se la scelta è basata solo sulla casualità? Qualcosa non quadra, ed in questo modo la casualità che ha retto la teoria stocastica fino ad ora è stata, almeno parzialmente, distrutta in un soffio.
L’olfatto che guida l’assone durante il lungo cammino
 L’ultima domanda è ‘Cosa succede con il collegamento al cervello quando forziamo una cellula ORC ad esprimere un recettore specifico scelto da noi?’. Gli esperimenti mostrano che il collegamento cervello-recettore è perfetto, quindi ne consegue che il collegamento è basato solo sull’OR espresso.
L’ultima domanda è ‘Cosa succede con il collegamento al cervello quando forziamo una cellula ORC ad esprimere un recettore specifico scelto da noi?’. Gli esperimenti mostrano che il collegamento cervello-recettore è perfetto, quindi ne consegue che il collegamento è basato solo sull’OR espresso.
Ma quando questa scelta cambia all’improvviso?
Per rispondere a questa domanda un gruppo di ricercatori ha tentato di cambiare il recettore scelto dalla cellula poco prima che questa riesca ad ‘agganciarsi’ al cervello (maturare), attraverso un gene OR esogeno ed inducibile.
In questo topo transgenico le ORC scelgono un recettore endogeno in un modo apparentemente casuale, e cominciano la propagazione dell’assone verso il cervello cercando la zona giusta per formare sinapsi. Durante questa fase si può innescare l’espressione del recettore transgenico con il conseguentemente spegnimento del recettore endogeno…
La cellula ancora immatura, come abbiamo visto, si ‘adatta’ al nuovo recettore, ma si aggancerà anche alla zona cerebrale giusta? La risposta è incredibilmente sì. il collegamento e la funzione del recettore è perfetto nonostante il cambio recettoriale durante la formazione dell’assone. Tuttavia, se si cerca di cambiare la scelta dell’OR espresso dopo la maturazione della ORC, si provoca una abberrazione che innesca apoptosi.
Come è possibile che un OR possa guidare la scelta della regione cerebrale giusta a cui ‘agganciarsi’ senza avere altri 1.000 geni deputati al riconoscimento dei diversi OR da parte del cervello?
Qualcuno ha suggerito un’ipotesi alquanto affascinante, secondo cui un OR si trovi alla testa dell’assone durante la fase di elongazione, guidandolo verso il corretto punto di aggancio al cervello come farebbe un cane da tartufo a portarci verso questo prezioso tubero in un intero bosco.
Teoria affascinante e stravagante, tuttavia ancora insoluta ed in cerca di qualcuno che la convalidi o la controbatta con esperimenti, o con ipotesi più semplici.
Conclusione
In questo lungo post abbiamo messo sul piatto tutto quello che è noto sull’olfatto senza uscirne fuori con una teoria solida in grado di spiegare tutti i dati sperimentali. Per chi fa ricerca questo risultato è il pane quotidiano poiché non si seguono mai strade battute, si cerca sempre di crearne nuove, con nuove ipotesi, nuove verifiche e nuove deduzioni che porteranno ad altre ipotesi che si spera siano migliori delle precedenti e più vicine alla verità.
Tutt’ora ci sono gruppi di ricerca che analizzano le seguenze di DNA dei recettori OR cercando qualcosa in comune; analisti in cerca di modifiche epigenetiche in questi clusters genomici; cacciatori di nuovi meccanismi cellulari; esperti di transgeni che tentano nuove e più affascinanti teorie, a volte anche bizzarre… e poi ci sono ricercatori che provano a dare un senso a questo puzzle di dati disarmonici. Il tutto per svelare un mistero che dura da tanti anni oramai.
Non vi è dubbio che ci sono persone che provano un certo fascino nel partecipare a questa continua caccia alla soluzione del problema strato dopo strato, tirando fuori teorie sempre più complesse ed affascinanti, ed a volte come in questo caso si tirano fuori addirittura nuovi meccanismi che si ignoravano fino a poco tempo fa.
Un in bocca al lupo a chi si addentrerà nel risolvere questo mistero anche solo per 5 minuti ed un grazie a Nico e Patrizio per l’incoraggiamento ed i consigli che mi hanno dato per la stesura di questo post.
Tags:
Animali transgenici,
Apoptosi,
Biologia molecolare,
Cellule Recettoriali,
Cre/LoxP,
fattori trascrizionali,
mappa sensoriale,
Neuroni,
neuroni olfattivi,
Olfatto,
Percezione
15 maggio 2009 - 12:03 pm
Rieccoci ancora per un’altra puntata per la divulgazione scientifica sull’ischemia cerebrale. Questo che vedremo è, in ordine cronologico, l’ultimo capitolo da me scritto ma il primo da me adattato per la pubblicazione di questa serie. Il motivo è che, a parer mio, potrebbe essere più intuitivo per le persone che non si sono mai affacciati ad un discorso scientifico impegnato. Trattasi di circa 15 anni di ricerche in questo campo seguito e presentato per lo più in ordine cronologico con qualche inesattezza o approssimazione in qualche punto per rendere il discorso quanto più scorrevole e divulgativo possibile; Vogliate scusarmi se in qualche caso ho semplificato un po’ troppo quanto si conosce realmente in letteratura.
Preambolo
Per chi non avesse seguito il percorso del nostro misterioso ‘giallo’ sull’ischemia cerebrale forse sarebbe bene leggersi i capitoli precedenti per poter seguire meglio questo ‘racconto’
http://www.molecularlab.it/insideneuroscience/?p=26
http://www.molecularlab.it/insideneuroscience/?p=37
http://www.molecularlab.it/insideneuroscience/?p=41
Siamo nelle vesti di un improbabile investigatore molecolare con il compito di risolvere un misterioso omicidio di cellule neuronali in seguito ad ictus. Fino ad ora abbiamo cercato di ricostruire la vita dei neuroni, le ultime ore delle vittime e come si presentano agli occhi della scena del crimine. In questo capitolo, invece, cercheremo di puntare il dito sul primo imputato eccellente dell’ischemia, ovvero un fattore trascrizionale ‘particolare’. Già la sola appartenenza di una proteina alla categoria “fattore trascrizionale” conferisce quell’alone di mistero ed ambiguità tipico, infatti, è noto che queste proteine frequentano “strane compagnie”, tale che il più amabile difensore della vitalità cellulare può trasformarsi nel peggior nemico da abbattere nelle neoplasie.
Ecco a voi quanto segue.
La prima pista delle indagini
 Una delle osservazioni più ovvie che mi sono state fatte nel tempo si basa sul fatto che tutto quello che si avvia durante l’ischemia cerebrale è senza alcun dubbio innescato dalla mancanza di ossigeno e nutrimento… secondo questa ipotesi ci si dovrebbe concentrare sugli eventi che sono attivati dall’ipossia e/o glucopenia per arrivare al vero meccanismo che innesca la morte neuronale ritardata. L’osservazione è ottima, e tantissimi ricercatori si sono concentrati su questa pista per trovare i responsabili.
Una delle osservazioni più ovvie che mi sono state fatte nel tempo si basa sul fatto che tutto quello che si avvia durante l’ischemia cerebrale è senza alcun dubbio innescato dalla mancanza di ossigeno e nutrimento… secondo questa ipotesi ci si dovrebbe concentrare sugli eventi che sono attivati dall’ipossia e/o glucopenia per arrivare al vero meccanismo che innesca la morte neuronale ritardata. L’osservazione è ottima, e tantissimi ricercatori si sono concentrati su questa pista per trovare i responsabili.
Sicuramente uno dei target più dibattuti in questo contesto è il fattore trascrizionale che porta il nome di Hypoxia Inducible Factor (HIF), ovvero fattore indotto dall’ipossia. Ce ne sono 3, chiamati HIF-1, HIF-2 e HIF-3; di questi solo il primo è noto alla maggioranza della comunità scientifica, e sembra essere il più importante.
Quale personaggio per HIF-1?
 Se dovessi associare questo fattore trascrizionale ad un personaggio di un giallo, potrebbe essere rappresentato senza alcun dubbio da un antipatico ed ambiguo individuo che attira su di sé tutti gli indizi, in modo tale che rappresenti l’assassino nell’immaginario del lettore per buona parte del racconto. In genere queste persone durante il racconto possono trovarsi alternativamente tra i principali sospetti, tra i fiancheggiatori, tra gli innocenti o tra i più improbabili difensori della verità.
Se dovessi associare questo fattore trascrizionale ad un personaggio di un giallo, potrebbe essere rappresentato senza alcun dubbio da un antipatico ed ambiguo individuo che attira su di sé tutti gli indizi, in modo tale che rappresenti l’assassino nell’immaginario del lettore per buona parte del racconto. In genere queste persone durante il racconto possono trovarsi alternativamente tra i principali sospetti, tra i fiancheggiatori, tra gli innocenti o tra i più improbabili difensori della verità.
Assassino, complice, arma, ancora di salvezza e persino futile perdita di energia… queste sono le definizioni che nel tempo sono state associate a questo strano fattore trascrizionale.
I Primi indizi ed il primo incontro
La pista dell’ipossia ci porta a chiedere se la scena del crimine era frequentata da fattori che sono proprio regolati da questa condizione. Da un elenco di proteine potremmo scorgere un personaggio molto influente definito HIF-1 e già qui giungiamo al primo mistero; dall’analisi della zona ischemica emerge chiaramente che HIF-1 non è presente al momento del ‘delitto neuronale’, un semplice Western Blot per l’analisi delle proteine mostra chiaramente una debolissima banda di HIF-1 ma niente di più di quello che si troverebbe in un qualsiasi tessuto in condizioni normali. Il personaggio quindi sembra non avere indizi a carico, tuttavia ci sono delle stranezze da puntualizzare.
Tiriamo fuori il nostro asso nella manica, ovvero la ricostruzione dell’avvenimento in un modello di ischemia su animale ed analizziamo la quantità di HIF-1 in diverse aree cerebrali a diversi tempi dopo l’occlusione dell’arteria cerebrale media. Dai risultati ci renderemo conto che HIF-1 non era presente prima dell’occlusione e neanche all’inizio dell’occlusione stessa, mentre è presente per pochissimo tempo durante l’ipossia neuronale per poi sparire di nuovo improvvisamente subito dopo il ripristino del circolo sanguigno.
HIF-1 dovrebbe spiegarci molte cose al riguardo sul perché c’era, cosa ha fatto e come ha fatto ad apparire e scomparire velocemente in circostanze quantomeno sospette.
Il mago Houdinì – Apparizioni e Sparizioni
HIF-1 è un fattore trascrizionale composto da due sub-unità definite HIF-1alfa e HIF-1beta, prodotti da due geni diversi molto conservati nell’evoluzione dei mammiferi. La trascrizione e la formazione di queste due proteine sono costanti ed ubiquitari in tutto l’organismo, tuttavia con un banale western blot non è possibile vedere una buona dose di proteina nei tessuti.
Il perché è subito chiarito dal suo scopritore Gregg Semenza, che ha dimostrato come in condizioni di ossigenazione ‘normale’ HIF-1alfa è idrossilato attraverso un enzima e poi processato per la degradazione… in poche parole HIF-1alfa è continuamente prodotto ma è anche degradato subito dopo in presenza di ossigeno molecolare. Infatti, quando l’ossigeno comincia a mancare HIF-1alfa non si può più degradare e si accumula legandosi al suo cofattore HIF-1beta formando il fattore trascrizionale completo HIF-1 che va ad attivare una serie di geni target. In seguito alla riossigenazione HIF-1alfa è nuovamente degradato ed il complesso HIF-1 non si forma più.
Ecco così spiegato il mistero della sparizione ed apparizione di HIF-1 durante l’ischemia cerebrale.
Conosci gli amici che frequenti e saprai chi sei
La domanda più interessante a questo punto è “quali geni target attiva HIF-1?”. Bhé ce ne sono centinaia, e sono divisi in classi secondo il meccanismo globale a cui afferiscono.
Troviamo geni del metabolismo anaerobico del glucosio, ovvero quegli enzimi che servono per ottenere energia dal glucosio senza la necessità di ossigeno, troviamo anche geni che servono per aumentare l’ossigenazione in una certa regione del tessuto come ad esempio VEGF il fattore che induce la vascolarizzazione in loco per apportare più ossigeno e nutrienti. Tra i target c’è anche l’eritropoietina che aumenta la produzione di globuli rossi che portano l’ossigeno in tutto l’organismo.
Poi ci sono geni un po’ più strani che mirano comunque alla sopravvivenza della cellula in condizioni anaerobiche e geni di cui non si sa bene la funzione nell’ipossia.
Tutto sommato è un ‘buono’ e serve alle cellule per ‘adattarsi’ alle scarse riserve di ossigeno e soprattutto promuovere il rimodellamento dell’organismo per avere più ossigeno.
Buono?
C’è anche l’altra faccia della medaglia, come per ogni fattore trascrizionale. L’altra faccia della ‘Janus Face’ HIF-1 è che se passa ‘dall’altra parte’ favorisce la vascolarizzazione di neoplasie (tumori) rendendolo più maligno e più resistente alle terapie antineoplastiche. Non a caso le mutazioni dei geni che servono per degradare HIF-1 in presenza di ossigeno siano dei noti oncogeni, ovvero geni cruciali che favoriscono la formazione e l’accrescimento dei tumori quando non funzionano a dovere.
Cosa ci faceva HIF-1 durante l’ischemia?
 Bene dopo questa velocissima presentazione di questo angelo o diavolo è ora di chiedersi cosa stava facendo subito dopo l’occlusione dell’arteria cerebrale e proprio in quei neuroni che moriranno diverse ore dopo… Stava avvelenando il loro futuro o stava costituendo un disperato tentativo di salvataggio?
Bene dopo questa velocissima presentazione di questo angelo o diavolo è ora di chiedersi cosa stava facendo subito dopo l’occlusione dell’arteria cerebrale e proprio in quei neuroni che moriranno diverse ore dopo… Stava avvelenando il loro futuro o stava costituendo un disperato tentativo di salvataggio?
Se provate a chiedere a HIF-1 cosa stesse facendo la risposta è ovvia, stava promovendo la sintesi di geni del metabolismo anaerobico per consentire ai neuroni di far fronte all’immediata assenza di ossigeno.
Facendo una Cromatin Immunoprecipitation (ChIP), ovvero una tecnica che ci permette di determinare se un fattore trascrizionale è legato o no ad una determinata regione genomica, c’è un primo riscontro positivo, effettivamente HIF-1 si trovava nelle posizioni di attivazione dei geni che servono per il metabolismo anaerobico.
Però c’è un’incongruenza, il tempo necessario ad HIF-1 per attivare i geni del metabolismo è costituito almeno da diverse ore; ovvero il tempo necessario per aumentare la sintesi di mRNA e di proteine. HIF-1, invece, rimane legato ai geni descritti per pochissimo tempo, al massimo un’ora o anche molto meno, un tempo che non basterebbe neanche a reclutare tutti i cofattori necessari per l’attivazione della trascrizione genica… come mai?
HIF-1 si difende esponendo i propri fatti… ovvero si trovava lì al momento dell’ipossia, aveva visto che ‘la nave stava affondando nell’ipossia’ e quindi si è attivato quanto prima come da suo dovere, poi è subentrata la riossigenazione dell’area cerebrale che lui non poteva prevedere ed è stato degradato dall’ossigeno prima che potesse fare effettivamente qualcosa. Per cui in realtà si tratterebbe di un tentativo andato a vuoto.
Dall’analisi per RT-PCR, tecnica che permette di determinare l’aumento o la diminuzione dell’espressione di determinati geni, c’è una prima conferma dell’alibi di HIF-1. Effettivamente stavano aumentando, seppur di poco, la sintesi dei geni implicati nel metabolismo anaerobico, del tutto sovrapponibile con quello che ci si aspetterebbe dall’azione esplicata da HIF-1.

Innocente allora?
Prima di prosciogliere l’imputato io suggerirei di analizzare anche chi ha frequentato HIF-1 in quella notte misteriosa dove è avvenuta l’ipossia. C’erano altri geni che ha indotto HIF-1?
Un gene molto importante è VEGF come già accennato, implicato nell’induzione della vascolarizzazione a lungo termine, ma che ha anche un’azione neuroprotettiva acuta nei confronti dell’ischemia cerebrale. Poi ci sono l’eritropoietina, che serve per aumentare i globuli rossi nel sangue, anch’esso neuroprotettivo in fase acuta, e tanti altri geni più o meno noti per avere un’azione neuroprotettiva. Un eroe dunque.

Il lato oscuro di HIF-1
C’è però anche chi fa notare che i fattori trascrizionali in condizioni non fisiologiche come l’ischemia cerebrale possono frequentare anche ‘compagnie diverse’ ed insolite da quelle catalogate. Infatti, da alcuni esperimenti di Microarray, tecnica che permette di vedere l’aumento o la diminuzione dell’espressione di un ampio numero di geni in condizioni fisiologiche rispetto a quelle patologiche, è emerso che HIF-1 ha scarsamente indotto geni neuroprotettivi, come già descritto, però durante la sua attivazione sono aumentati enormemente anche tanti geni di ‘malaffare’.
Insomma tanto basta per iniziare il processo contro questo ‘strano individuo’
Che il processo abbia inizio… entri il primo imputato HIF-1
 La parola all’accusa
La parola all’accusa
Quei pochi minuti che bastano
Sebbene siano necessarie diverse ore per attivare i geni del metabolismo anaerobico, per HIF-1 sono stati necessari pochi minuti per attivare alcuni dei geni più distruttivi dell’ischemia. E non si tratta di una piccolo aumento di espressione. Il gene in questione è la nitrossido sintetasi inducibile che produce radicali liberi sottoforma di gas nitrossido con un’azione neurotossica letale per i neuroni. Da un’analisi con Decoy, ovvero la somministrazione di piccoli tratti di DNA a doppio filamento che simulano il sito di legame di HIF-1 e lo sequestrano lontano dai geni bersaglio, è stato possibile determinare con certezza che HIF-1 è il principale responsabile dell’aumento della trascrizione della iNOS. Quando ad una cellula ipossica si dà il Decoy, HIF-1 lo lega e non riesce a raggiungere il gene bersaglio, un effetto simile ad un knock-out genico. La iNOS è nota per causare la morte cellulare dei neuroni, una questione indubbia.
Ma come è possibile che ad HIF-1 bastino pochi minuti per l’attivazione di iNOS mentre sono necessarie ore per i geni neuroprotettivi? Greeg Semenza è riuscito a chiarire anche questo altro aspetto. HIF-1, infatti, è prodotto continuamente dalla cellula, ed è anche degradato continuamente, ma una piccola quota di HIF-1 è presente nelle cellule, visibile per western blot, e basta per legare ed indurre determinati geni insoliti. Talvolta li lega anche quando non c’è ipossia ed attende la mancanza di ossigeno così per trovarsi già in loco quando serve.
Insomma HIF-1 è stato visto aumentare e fomentare geni per la degenerazione neuronale, mentre i geni neuroprotettivi non erano stati trattati allo stesso modo, dando solo un’apparenza di eroe salvatore.
A carico di HIF-1 c’è anche la precisione di espressione proprio nei neuroni e nei luoghi dove ci sarà, diverse ore la sua scomparsa, la maggior percentuale di neuroni morti. Una coincidenza forse?
Insomma, la situazione si è messa male per HIF-1, le prove vanno decisamente contro; il legame degli enzimi metabolici e neuroprotettivi potrebbe essere solo una copertura per il vero scopo, ovvero la degenerazione dei neuroni?
La parola alla difesa
 La difesa tira fuori il dogma scientifico
La difesa tira fuori il dogma scientifico
Il Dogma scientifico
Per chi non lo sapesse, esiste un dogma per ogni disciplina culturale umana e la biologia non ne è immune. Il Dogma dei biologi è che niente succede a caso, poiché per ogni gene o proteina ci sono milioni di anni di evoluzione che raffina tutti i meccanismi a quelli fondamentali per la sopravvivenza e per l’evoluzione. Se ci fosse qualcosa di inutile o peggiorativo sarebbe eliminato automaticamente dall’evoluzione stessa. Per cui la selezione e la conservazione di un fattore indotto dall’ipossia che causi morte nei neuroni non è credibile poiché non darebbe un vantaggio per la sopravvivenza.
Dopotutto quello che si vede è una sommatoria di tantissimi meccanismi che interagiscono tra di loro e quello che si osserva è solo la summa del tutto da cui è difficile estrapolare un singolo messaggio. Meglio ricorrere all’incidente probatorio…
Ovvero spegniamo HIF-1 con un knock-out genico ed induciamo l’ischemia, verifichiamo in questo caso se l’animale che manca di questo fattore è più o meno esposto al danno neuronale rispetto agli animali wild-type.
L’incidente probatorio o la prova del nove
La prova più importante seguita da quella definitiva molto spesso è la generazione di un topo knock-out per il gene di interesse. Ci sono stati tantissimi esempi di questo tipo, vale sia come inizio di una ricerca per capire grossolanamente un gene di cosa sia responsabile e vale anche come prova raffinata per avere la prova definitiva che un determinato gene sia coinvolto in un certo meccanismo.
Purtroppo nonostante intensi sforzi per ottenere topi knock-out per HIF-1alfa, è emerso che un animale privato di questo gene non riesce a svilupparsi ed a sopravvivere ai primi stadi embrionali, anche se è esposto ad un normale livello di ossigenazione. Ovviamente questo è stato il motivo dei ritardi nel fare un topo transgenico e quindi definire una prova evidente del suo ruolo in tanti processi fisiologici e patologici.
Tuttavia, recentemente è stato possibile generare un topo con un knock-out tessuto specifico di HIF-1alfa per i neuroni, il risultato è stato quantomeno sorprendente… l’assenza di HIF-1alfa durante l’ischemia cerebrale ha enormemente potenziato i danni neuronali.
Un fulmine a ciel sereno per tutti… L’imputato è senza dubbio da scagionare e da mettere tra i difensori cellulari.
Conclusioni
In questo capitolo abbiamo visto un esempio di imputato eccellente dell’ischemia cerebrale che ancora oggi per alcuni gruppi di ricerca rappresenta ancora il centro dell’attenzione. Gli studi andranno sicuramente avanti, anche se non ci si aspetterebbe ancora molto da quanto descritto, HIF-1alfa nel processo dell’ischemia cerebrale si potrebbe definire almeno non pericoloso tanto quanto altri imputati eccellenti che vedremo nelle prossime puntate. Per questi motivi l’attenzione su questo fattore trascrizionale si è concentrato soprattutto sulle neoplasie, ed ha tralasciato progressivamente l’ischemia cerebrale, renale e cardiaca. Vedremo se qualcuno in futuro troverà qualche prova rilevante per riaprire nuovamente il processo. In attesa di tale data ci rivedremo nella prossima puntata dove vedremo un altro imputato importante.
Tags: fattori trascrizionali, HIF-1, HIF-1alfa, Ictus, Ischemia Cerebrale, Mente, Neuroscienze
5 gennaio 2009 - 11:52 am
Preambolo
Carissimi,
recentemente mi è capitato di incontrare una patologia rara, che ignoravo fino a poco tempo fa, e che ha innescato in me pensieri e teorie contrastanti a cui non saprei dare un giusto ordine.
Per capirci, come al solito, c’è bisogno di fare un po’ d’ordine nel linguaggio. Il punto d’origine dei nostri assi cartesiani verbali in questo post sarà la parola ‘Normale’ che identifica, senza ombra di dubbio, una qualsiasi caratteristica che si presenta nella quasi totalità della popolazione. Da questo ‘punto zero’ potremmo allontanarci definendo un maggiore o minore grado di ‘anormalità’. Si potrebbe persino tracciare un asse con una direzione per ‘anormalità buone’ e ‘cattive’ e deporre in queste due zone tutte le possibili caratteristiche che ci fanno allontanare dalla normalità. Questa semplificazione, purtroppo, ci porta ad un limite, cioè per quanto ci potremmo sforzare alcune ‘anormalità’ potrebbero essere difficili da sistemare. Per intenderci, tutti noi saremmo certamente lieti di essere ‘anormalmente intelligenti e forti’, molto meno per ‘anormalmente bassi e brutti’. Tuttavia, alcune di queste certezze sarebbero perse quando dovremmo ipoteticamente posizionare in questo contesto la sindrome di Asperger.
Cosa è la Sindrome di Asperger?
 Cosa è la Sindrome di Asperger?
Cosa è la Sindrome di Asperger?
Si potrebbe definire come un innato disordine del comportamento, ma è difficile avere una definizione di questa patologia in due parole, forse potrebbe essere meglio considerata semplicemente come un insieme di sintomi con una maggiore o minore gravità.
Se vi dicessi che alcuni studiosi pensano che Michelangelo, Wittgenstein, Kurt Gödel, Glenn Gould, Bobby Fischer, Satoshi Tajiri, The Vines Craig Nicholls, Albert Einstein e Isaac Newton soffrissero in qualche misura di questa ‘sindrome anormale’ e che proprio questa caratteristica donasse loro un qualcosa in più rispetto alla ‘normalità’?
Se tutto questo fosse vero, lo stesso concetto di ‘malattia’ o ‘sindrome’ si rivaluterebbe in qualcosa di assolutamente meraviglioso, anzi molti si precipiterebbero ad informarsi su come potersi ‘ammalare’ di questa ‘anormalità’.
L’anormalità è quindi positiva? Come ho già accennato non si tratta di una cosa semplice, poiché la malattia in questione è la sindrome di Asperger, un disordine psicologico che secondo alcuni appartiene alle mille sfaccettature dell’autismo. Già il concetto di autismo è di difficile comprensione, e la sindrome di Asperger sembra esserne una ulteriore complicazione, poiché prende alcune delle peggiori caratteristiche dell’autismo con le più belle caratteristiche che l’umanità possa desiderare in una complessa macchina psicologica completamente impenetrabile ad una semplice analisi.
Vi prego di non immaginare che le persone affette da questa ‘anormalità’ appaiono chiuse e taciturne come Dustin Hoffman in Rain Man; tutt’altro, la loro facilità di linguaggio è tale da poter essere definiti “iperlessici”. Si può dire anche che hanno un raro e sviluppato senso dell’umorismo, con un’abilità non comune per capire e creare doppi sensi, giochi linguistici, satira ed altro. Le persone con la sindrome di Asperger, inoltre, appaiono come delle persone molto intelligenti ed in molti casi dei veri e propri geni. Sono immersi nella società e spesso fanno dei lavori molto impegnativi e che richiedono una particolare intelligenza, cultura o concentrazione; ad esempio architettura, ingegneria, programmazione informatica e persino la chirurgia medica. Persone abbastanza ‘normali’ per trovarsi immersi nella società, ma abbastanza speciali per non essere classificati come persone mediocri anche se spesso non si rendono conto di essere così speciali.
I soggetti ‘malati’ della sindrome di Asperger sono spesso attratti, con un intenso livello di attenzione, solo da determinate cose che interessano loro e da attività in cui si possa ritrovare un certo ordine come le classificazioni, liste e simili.
L’individuo può portare avanti anche una vita di successo, poiché spesso manifesta ragionamenti estremamente sofisticati, un’attenzione pressoché ossessiva e una memoria eidetica, ovvero focalizzata sulle immagini visuali e sui dettagli, utile ad esempio nella chirurgia.
Ma allora cos’è che manca? Perché si definisce come sindrome?
L’altra faccia della medaglia
 Ovviamente tutto ha un prezzo, e queste persone pagano un prezzo altissimo per queste straordinarie ‘anormalità’. Quello che manca a loro è un’innata abilità di capire ed esprimere gli stati emotivi con gesti, linguaggio corporeo ed espressione facciale.
Ovviamente tutto ha un prezzo, e queste persone pagano un prezzo altissimo per queste straordinarie ‘anormalità’. Quello che manca a loro è un’innata abilità di capire ed esprimere gli stati emotivi con gesti, linguaggio corporeo ed espressione facciale.
Le persone affette da sindrome di Asperger, sebbene non manchino di intelligenza e capacità linguistiche, appaiono incapaci di comprendere il significato di semplici segnali come un sorriso, che magari può essere interpretato come una semplice smorfia più che un segno di gradimento. Mancano, inoltre, dell’abilità di capire ciò che non viene detto esplicitamente ed in pari modo essi hanno difficoltà a comunicare con accuratezza il loro proprio stato emotivo.
Mentre la maggioranza delle persone al mondo si trova più o meno nella media per qualunque caratteristica fisica o psichica e di conseguenza non brilla per nessuna di queste, i bambini affetti da questa strana sindrome, invece, brillano come ‘piccoli professori’ in materie a loro piacevoli da studiare e spesso potrebbero mettere in difficoltà anche persone di livello universitario su determinati argomenti per cultura, profondità di pensiero e spiccato senso critico. Tutta questa straordinaria capacità di concentrazione, intelligenza, genialità, cultura, passione per lo studio e per la ricerca, però, è impotente di fronte alla capacità di percepire gli stati mentali degli altri a livello cognitivo ed emotivo. Le persone con la sindrome di Asperger possono osservare un sorriso e non capire se si tratti di un segno di comprensione, di accondiscendenza o di malizia, e nei casi più gravi non riescono neppure a distinguere la differenza tra sorriso, ammiccamento e altre espressioni non-verbali di comunicazione interpersonale. Per loro è estremamente difficile saper “leggere attraverso le righe”, ovvero capire quello che una persona afferma implicitamente senza dirlo direttamente.
Bisogna comunque notare che, trattandosi di un disturbo con ampio spettro di variazione, una certa percentuale di individui con la sindrome di Asperger appaiono quasi normali nella loro capacità di leggere le espressioni facciali e le intenzioni degli altri. Molti di questi inconsapevoli geni, infatti, non sanno neanche di essere affetto da una forma lieve di questa patologia, mentre altri più gravi mostrano difficoltà anche nel guardare negli occhi le altre persone, ritenendolo pericoloso e al di sopra delle proprie possibilità, oppure hanno un contatto visivo eccessivamente fisso, che può essere avvertito come “disturbante” per le persone comuni.
Genialità: Croce e Delizia
 Bisogna parlare anche della parte più oscura della sindrome di Asperger che come avrete intuito non dà quello che si potrebbe definire una vita tutta rose e fiori. Spesso può portare al soggetto molti problemi nelle interazioni sociali tra persone di pari livello. Un bambino affetto da tale sindrome, ad esempio, ha difficoltà a decodificare i segnali impliciti su cui si reggono le interazioni sociali e può essere messo da parte dai coetanei, generando un’alienazione sociale talmente intensa che il bambino può arrivare a creare degli amici immaginari per sentirsi in compagnia.
Bisogna parlare anche della parte più oscura della sindrome di Asperger che come avrete intuito non dà quello che si potrebbe definire una vita tutta rose e fiori. Spesso può portare al soggetto molti problemi nelle interazioni sociali tra persone di pari livello. Un bambino affetto da tale sindrome, ad esempio, ha difficoltà a decodificare i segnali impliciti su cui si reggono le interazioni sociali e può essere messo da parte dai coetanei, generando un’alienazione sociale talmente intensa che il bambino può arrivare a creare degli amici immaginari per sentirsi in compagnia.
I problemi proseguono anche nella vita adulta, molti individui con la sindrome di Asperger, infatti, non possono avere una vita considerata socialmente appagante dalla gente comune, molti infatti rimangono pressoché soli, anche se è possibile che stringano strette relazioni con alcuni individui.
Stiamo trattando di una patologia con un diverso grado di severità, quindi si troveranno anche molte persone, che soffrendo di questo disturbo, si sono sposate e hanno avuto figli; i loro bambini possono essere persone comuni o soffrire di qualche disturbo di tipo autistico.
Tutt’oggi la causa della sindrome di Asperger non è ancora nota, anche se si sa che c’è una forte influenza genetica e non ci sono differenze neurologiche confrontati ai soggetti così detti ‘normali’.
I soggetti non hanno avuto alcun trauma evidente da piccoli, non sviluppano un ritardo nel parlare e non manca il desiderio di comunicare con l’esterno.
Tutto sommato le persone con la sindrome di Asperger sono vittime inconsapevoli dei lati oscuri della genialità, sono più facili a cadere in una forma depressiva rispetto alla popolazione generale perché essi hanno spesso difficoltà a comunicare problemi o capire quando è il momento di mostrare affetto. Amano come gli individui ‘normali’, e forse di più perché spesso sono soli, ma sono molto letterali nel parlare e hanno difficoltà nel comunicare in maniera emozionale, e frequentemente possono instillare nella propria compagna insicurezza e confusione.
Riflessioni
 Malattia quindi? o dono divino? Un fardello, appesantito dalla solitudine, da portare sulle spalle o semplici effetti collaterali della genialità?
Malattia quindi? o dono divino? Un fardello, appesantito dalla solitudine, da portare sulle spalle o semplici effetti collaterali della genialità?
Sicuramente le persone con la sindrome di Asperger non si potrebbero definire come semplici pazienti da ‘curare’, ma persone con grandi doti di intelligenza che possono contribuire attivamente allo sviluppo della società e della civiltà, come è già successo in passato. Il trattamento per la sindrome di Asperger, quindi, non c’è, ma attualmente si cerca di migliorare gli effetti collaterali che questi sintomi possono procurare. Poiché in fondo, spesso sono persone che hanno solo bisogno di essere accettati per le proprie caratteristiche. Come un non vedente che vuole dalla società un mondo più accessibile per non esserne escluso… allo stesso modo c’è chi non legge le espressioni degli altri e vorrebbe la stessa cosa. Immaginate chi si dovrebbe definire portatore di handicap… Un genio che ‘non vede’ le espressioni del viso? Oppure le persone ‘normali’ che hanno un intelletto più limitato?
Forse in questo caso la definizione ‘diversamente abile’ sarebbe più azzeccata che mai.
Tags: anormalità, autismo, genialità, handicap, Mente, Neuroscienze, normalità, Percezione, Psicologia, Sindrome di Asperger, Visione
17 dicembre 2008 - 9:55 pm
La ricostruzione della scena del crimine
 Allora, per chi non avesse seguito le puntate precedenti, abbiamo introdotto un giallo misterioso dove delle cellule del cervello umano muoiono improvvisamente in seguito ad ischemia cerebrale (icutus), mentre noi siamo la polizia scientifica molecolare nominati per cercare di spiegare cosa sia accaduto in quegli istanti e determinare chi ha causato la morte neuronale.
Allora, per chi non avesse seguito le puntate precedenti, abbiamo introdotto un giallo misterioso dove delle cellule del cervello umano muoiono improvvisamente in seguito ad ischemia cerebrale (icutus), mentre noi siamo la polizia scientifica molecolare nominati per cercare di spiegare cosa sia accaduto in quegli istanti e determinare chi ha causato la morte neuronale.
Per chi non avesse seguito le parti precedenti dia uno sguardo ad i seguenti links Ischemia Cerebrale (Parte I) Ischemia Cerebrale (Parte II)
Fino ad ora il nostro taccuino è quasi vuoto, sappiamo ancora poco e ci sono ancora tante cose da analizzare. Se fossimo degli zelanti ispettori, una delle prime cose da fare potrebbe essere ricostruire le ultime ore delle vittime per capire chi poteva trovarsi là al momento del delitto. In questo caso potremmo usare dei modelli cellulari (in vitro) o animali (in vivo) per riprodurre quello che succede nell’ictus umano. La scelta del modello non è banale, anzi è fondamentale per poter chiarire qualcosa. In genere si hanno due strade davanti ed in base a questa scelta dipenderà il livello di difficoltà che si dovrà affrontare.
La prima strada è la scelta di modelli cellulari (in vitro) che danno dei dati semplicemente misurabili ed abbastanza riproducibili; tutto sommato è un sistema semplicissimo da affrontare, ma d’altra parte ci accorgeremo presto che questi modelli sono un’eccessiva semplificazione di quello che avviene nell’uomo e che quindi ci sono anche delle notevoli limitazioni.
La seconda strada, invece, è la scelta di un modello animale (in vivo) che riproduce con molta fedeltà quello che avviene sull’uomo, ma è difficile da capire ed analizzare in tutti i suoi dettagli, proprio perché più simili al danno umano.
Ci sarebbe anche una terza strada definita ex vivo oppure colture organo tipiche, in cui si coltivano interi tessuti o parti di organi prelevati dall’animale vivo per avere una maggiore complessità rispetto alle semplici cellule, ma un vantaggio di semplificazione rispetto all’animale intero.
Le ultime ore delle vittime
Quello che si fa è prendere un certo numero di animali, ad esempio topi o ratti, ed indurre in questi l’ictus attraverso l’occlusione di una arteria cerebrale importante. Ogni animale potrebbe essere analizzato in tempi diversi così da ricostruire gli eventi che seguono l’occlusione sanguigna e precedono il danno finale.
I risultati sono interessanti, poiché è subito evidente che non tutte le cellule neuronali muoiono contemporaneamente; il volume ischemico si espande da una zona più vicina all’occlusione sanguigna fino ad arrivare ad un massimo di espansione a circa 24 ore dal danno. In ogni momento potremmo riconoscere una zona prevalentemente necrotica con poche cellule vive (core ischemico) ed una zona circostante in sofferenza che diverrà ben presto anch’essa morta (penombra ischemica).
L’onda killer
 Se dovessimo ricostruire cosa accade negli ultimi istanti di queste cellule, ci potremmo accorgere che le cellule della penombra moriranno entro qualche ora non riescono a gestire i flussi di calcio citosolico (vedi primo post). Il calcio si accumula nelle cellule poiché queste non hanno energia a sufficienza per pomparlo fuori; di conseguenza le cellule sono più facilmente eccitabili e possono attivarsi anche in momenti non opportuni, liberando neurotrasmettitori che eccitano altre cellule come un’onda di scariche elettriche longitudinali che si propagano dal centro dell’ischemia fino alla periferia del cervello. Proprio in questi momenti, ovvero poco prima della morte cellulare, si possono avere dei corto circuiti neuronali e l’avvio di sistemi di ripristino di emergenza nella metà cerebrale colpita. Questi corto circuiti generano i classici segni premonitori che si possono riassumere nell’acronimo inglese FAST
Se dovessimo ricostruire cosa accade negli ultimi istanti di queste cellule, ci potremmo accorgere che le cellule della penombra moriranno entro qualche ora non riescono a gestire i flussi di calcio citosolico (vedi primo post). Il calcio si accumula nelle cellule poiché queste non hanno energia a sufficienza per pomparlo fuori; di conseguenza le cellule sono più facilmente eccitabili e possono attivarsi anche in momenti non opportuni, liberando neurotrasmettitori che eccitano altre cellule come un’onda di scariche elettriche longitudinali che si propagano dal centro dell’ischemia fino alla periferia del cervello. Proprio in questi momenti, ovvero poco prima della morte cellulare, si possono avere dei corto circuiti neuronali e l’avvio di sistemi di ripristino di emergenza nella metà cerebrale colpita. Questi corto circuiti generano i classici segni premonitori che si possono riassumere nell’acronimo inglese FAST
- Faccia asimmetrica (ad es. una palpebra più bassa, o una bocca storta): ciascuna metà del nostro organismo è gestita da una metà cerebrale, se in una di queste ultime avvengono corto circuiti oppure entrano in funzione sistemi di emergenza, la faccia o il corpo può avere un’asimmetria.
- Arti asimmetrici: si sollevano entrambe le braccia con i palmi rivolti verso l’alto, poi si chiudono gli occhi e si mantiene questa posizione per qualche secondo, nell’ictus un braccio tende a scendere per lo stesso motivo precedente
- Suoni confusi o distorti: Se le scariche elettriche colpiscono il centro del linguaggio, il paziente potrebbe essere colto da afasia con emissione di parole confuse, suoni distorti o potrebbe pensare di parlare correttamente mentre farfuglia parole a caso.
- Tempo: se si notano anche uno solo dei sintomi precedenti non bisogna esitare, una cura tempestiva è fondamentale per una buona riuscita della terapia.
Questi sintomi, infatti, compaiono ben prima che avvenga la morte dei neuroni, quindi sono ancora del tutto reversibili se si riesce a ripristinare la circolazione cerebrale.
Il paziente potrebbe anche avere all’improvviso
- debolezza o insensibilità o comparsa di formicolii a una metà del volto, a un braccio o a una gamba oppure a tutta la metà del corpo
- oscuramento o perdita della visione da un solo occhio
- incapacità di esprimersi o di comprendere qualcuno che parla
- inspiegabile sensazione di vertigine, capogiro, sbandamento o cadute improvvise
- grave e inspiegabile mal di testa.
Se questi sintomi scompaiono entro 24 ore, al paziente verrà detto che ha avuto un Transient Ischemic Attack (TIA), cioè un attacco ischemico transitorio, in pratica un ictus minore, dovuto a una occlusione solo transitoria di un vaso arterioso.
Primo intervento
Se il paziente colpito da icuts ha la fortuna avere vicino qualcuno che sa riconoscere i sintomi e riesce a portarlo in pronto soccorso entro le 3 ore dall’occlusione dell’arteria cerebrale, allora i medici potrebbero somministrare al paziente del tPA, ovvero l’attivatore tissutale del plasminogeno, in grado di degradare il trombo che ha occluso un’arteria e ridare ossigeno alle cellule neuronali prima che sia troppo tardi. Altra alternativa altrettanto valida è inserire un palloncino nell’arteria occlusa per gonfiarlo nell’ostruzione e liberare il circolo sanguigno.
Quando invece è già troppo tardi succede un qualcosa di irreversibile o quasi. Questo impercettibile punto di non ritorno che avviene dopo le prime 3 ore nell’uomo e ben prima che incominci la morte neuronale è l’oggetto della nostra contesa.
Fino ad oggi, infatti, non esistono metodologie che riescano in qualche modo ad identificare il perché le cellule neuronali muoiano anche se ricevono ossigeno, metaboliti e sostanze trofiche, e non si riesce a capire neanche quanto sarà esteso il danno, quali cellule moriranno e soprattutto quale zone saranno interessate. Ai familiari del paziente, infatti, verrà detto che bisognerà aspettare 1-2 giorni per capire quanto esteso è il danno e quali zone del cervello interesserà.
Questo aspetto è il punto cruciale, in queste ore che precedono il danno le cellule sono vive e ci servirebbe capire perché muoiono per poi cercare disperatamente un metodo per bloccare tale procedura apparentemente irreversibile.
Analisi della scena del crimine
 Un ispettore che si trova in una scena del crimine molto complessa come quella descritta e che ha sentito la ricostruzione del danno analizzerebbe sicuramente tutta una batteria di proteine e di mRNA per capire costa stavano facendo i neuroni in quei momenti. Questa analisi ovviamente può essere fatta da una ipotetica polizia scientifica utilizzando, ad esempio, la metodica dei microarray.
Un ispettore che si trova in una scena del crimine molto complessa come quella descritta e che ha sentito la ricostruzione del danno analizzerebbe sicuramente tutta una batteria di proteine e di mRNA per capire costa stavano facendo i neuroni in quei momenti. Questa analisi ovviamente può essere fatta da una ipotetica polizia scientifica utilizzando, ad esempio, la metodica dei microarray.
I microarray sono dei piccoli chips della grandezza della punta del nostro indice con cui è possibile valutare l’aumento o la diminuzione dell’espressione dell’mRNA di tantissimi geni contemporaneamente attraverso l’uso di un estratto di RNA da un controllo ed un estratto di RNA da un trattato.
Per chi non conosce i chips può leggere qui Microarray – Wikipedia
Tecnicamente questa metodica ci consente di guardare sommariamente tra i nostri 30.000 geni, quali sono aumentati o diminuiti in seguito ad un determinato stimolo, farmaco, malattia etc etc.
Questa metodica, infatti, si addice benissimo ad avere una visione globale del problema per incominciare un percorso di indagine sui responsabili della patologia.
Noi potremmo utilizzare questa metodica per analizzare zone analoghe del cervello in un animale ischemico e confrontarlo con un animale shame-operated, ovvero un animale che ha ricevuto una procedura chirurgica simile ma che non ha avuto l’occlusione di un arteria. Questo è importante poiché dobbiamo evitare alcune interferenze dovute allo stress operatorio, ad eventuali infiammazioni o infezioni. Dall’analisi emerge che sono variati almeno 7.000 geni.
Abbiamo quindi 7.000 testimoni o potenziali imputati da valutare con attenzione singolarmente o in gruppo nel processo che si sta aprendo, a cui si dovrebbero aggiungere anche altri imputati eccellenti che non possono essere valutati con i microarray, ad esempio neurotrasmettitori, sostanze endogene, ioni, meccanismi post-traduzionali (su proteine) che possono alterare l’espressione di una proteina, e la sua funzione senza interferire in alcun modo sulla sua espressione.
Conclusione
In questo ultimo post introduttivo abbiamo presentato un piccolo bagliore della difficoltà di questo lungo ed inesauribile processo verso ignoti che causano ogni anno una invalidità più o meno grave fino alla morte di tantissime persone nei paesi industrializzati. Per utilizzare un termine moderno si potrebbe parlare di un maxi-processo in cui gli imputati sono tra loro correlate con gerarchie complesse. Come già accennato, gli imputati sono in realtà numericamente molti di più di 7.000, ma fortunatamente non li prenderemo in considerazione tutti nel nostro blog; vedremo solo alcuni personaggi perché interessanti, ambigui, fortemente sospettati, antipatici, schivi o semplicemente perché si trovava lì al momento del delitto.
Alla prossima puntata.
Tags:
Ictus,
Ischemia Cerebrale,
Microarray,
mRNA,
Neuroni
14 settembre 2008 - 3:50 pm
Preambolo
Allora, rieccoci qui dopo le tanto meritate vacanze. Ringrazio tutti quelli che hanno contribuito con commenti, domande e quant’altro al primo capitolo di questa serie, spero che ce ne siano sempre di più, comprese critiche e puntualizzazioni; accetto persino complimenti.
Nel capitolo precedente abbiamo visto un po’ in generale alcune caratteristiche dell’ictus ed abbiamo introdotto il concetto del calcio come strumento di informazione e distruzione cellulare. Ora, in questo capitolo ci concentreremo maggiormente sulla morte neuronale, trattando il tutto come il più classico dei gialli dove le vittime sono i neuroni, la scena del crimine è il cervello dopo l’ictus e dovremo cercare l’assassino capitolo dopo capitolo, strato dopo strato, mettendo sul banco degli imputati diversi “personaggi ambigui” in cerca del vero colpevole. Di volta in volta saranno presentati nuovi personaggi con nuove “verità” e quindi nuove “teorie”. Ovviamente non mancheranno i colpi di scena come nel più classico dei thriller da leggere la sera a letto prima di dormire.
Data l’importanza dei rudimenti medici che stiamo trattando e dell’interesse mostrato in alcuni commenti ho deciso di aggiungere un piccolo paragrafo di chiarimenti e risposte ai commenti nel post precedente. Per chi si annoia a leggere le osservazioni fatte o per chi sa già questi concetti consiglio di saltare il prossimo paragrafo e passare direttamente al paragrafo sulla morte cellulare che spero sia per loro più interessante.
Il Calcio presente nel nostro organismo

Il calcio (simbolo: Ca2+) in una persona di sesso maschile magro rappresenta circa l’1,5% del peso corporeo, per una persona di 70 kg ce n’è circa 1 kg. Sicuramente è lo ione più abbondante del nostro organismo, seguito dal potassio (simbolo: K+) con soli 240 grammi e poi dal sodio (simbolo: Na+) con 100 grammi. Il 99% del calcio del nostro organismo è presente nelle ossa e nei denti, lo 0,5% è conservato avidamente in particolari compartimenti subcellulari come il reticolo endoplasmatico, insieme a proteine chelanti e mitocondri, come già descritto da Massi-Demish. Solo lo 0,3-0,4% del peso corporeo, circa 0,21 grammi, corrisponde al calcio che è libero di circolare nel nostro organismo e che partecipa a quei meccanismi di cui abbiamo parlato nel post precedente. Per dare un’idea, basti pensare che una mozzarella da 100 grammi contiene 0,4 grammi di calcio, ovvero una quantità doppia rispetto a tutto il calcio libero che è presente nel nostro organismo. Cosa succede se si assume una quantità di calcio superiore alle necessità dell’organismo? quasi sempre alcune cellule specializzate lo depositato nelle ossa oppure lo elimina attraverso le urine. Nei rari casi in cui c’è una carenza di calcio dalla dieta, invece, altre cellule specializzate corrodono meno di un millimetro cubo di ossa (quasi niente) per avere mesi di autonomia da calcio allo stato libero utile per i neuroni e tutte le altre funzioni cellulari descritte. Le nostre ossa sono infatti continuamente corrose e rimodellate in funzione del nostro lavoro fisico, delle trazioni, dai livelli ormonali e tante altre cose, di conseguenza l’organismo può sempre bilanciare l’eccesso o la carenza di calcio libero nei nostri fluidi corporei depositandolo o prelevandolo dalle ossa. Da questo si deduce che la quantità di calcio assunto per via orale di per sé non ha alcuna interferenza sulla quantità di calcio libero nel nostro organismo e di conseguenza non ha alcun effetto neanche sul danno ischemico.
Anche se si considerano i rari casi in cui delle persone, per qualche motivo, hanno una dieta cronica quasi del tutto priva di calcio e che quindi necessitano una integrazione, i livello di calcio libero sono più o meno simili alla media standard. A questi pazienti, però, può essere consigliato l’assunzione di pillole contenenti calcio oppure assumere qualche bicchiere di latte, pezzi di formaggio, carne, acqua del rubinetto per poter ripristinare le riserve di calcio nelle ossa ed evitare spiacevoli facili fratture.
C’è da dire anche che esistono, con una maggiore frequenza, delle persone che hanno carenze di calcio perché non lo riescono ad assumere dal cibo o lo elimina troppo velocemente, come nel caso dell’osteoporosi da menopausa. In questi casi si prescrivono farmaci composti da ormoni che predispongono l’organismo ad accumulare calcio nelle ossa anziché eliminarlo, senza la necessità di variare in alcun modo la dieta. Proprio questi ormoni influiscono sull’ischemia cerebrale, ma per meccanismi che non hanno nulla a che fare con la quantità di calcio ingerito, se volete posso aggiungere un capitolo apposito per questo curioso argomento.
Come può morire una cellula?

Allora, ora prendiamoci una pausa ed immaginiamo per un attimo di trovarci in un insolito thriller e di essere chiamati nel cuore della notte per investigare su un misterioso assassinio di massa. Una volta arrivati sulla scena del delitto, ovvero il cervello ischemico, ci ritroveremmo sicuramente ad analizzare come si presentano le vittime neuronali. Però c’è un problema, anche se fino ad ora abbiamo parlato intuitivamente di morte cellulare, non sappiamo ancora come può morire una cellula e di conseguenza come può presentarsi agli occhi di una insolita polizia scientifica molecolare. Questo è un punto fondamentale poiché ci aiuta a capire meglio la filosofia e la strada che stiamo percorrendo.
Attualmente si classifica la morte cellulare in due tipologie principali:
- Necrosi: generalmente è il risultato di un acuto stress o trauma che porta alla rottura della membrana cellulare con perdita del materiale biologico in esso contenuto. In questo caso la cellula si presenta in maniera inconsistente e quasi irriconoscibile poiché tutti gli organelli di cui è composta si disperdono nella soluzione in cui è immersa la cellula morta, rendendo spesso i resti irriconoscibili.
- Apoptosi: si tratta di un processo di morte molto ordinato in cui è la stessa cellula a decidere come e quando morire, e lo fa in modo tale da non dare alcuna tossicità alle cellule circostanti. La cellula si presenta come piccole vescicole ricoperte di membrana e con dei segnali sulla membrana cellulare (epitopi) che indicano “qui c’è materiale riciclabile”.
La necrosi, quindi, è insieme di morti cellulari “accidentali” senza avere un comune denominatore se non la rottura irreversibile della membrana che avvolge le cellule e di conseguenza la dispersione del materiale in esso contenuto. Proprio la perdita di materiale ed il disordine caratteristico di questo tipo di morte può causare infiammazione o tossicità alle cellule circostanti. Sicuramente la necrosi non è un fenomeno che l’organismo e le cellule stesse apprezzano o considerano come destino. L’apoptosi invece è una forma di morte cellulare molto precisa, portata avanti in modo ordinato e regolato, che dà un vantaggio alla popolazione cellulare circostante durante il ciclo vitale dell’organismo. Si tratta in realtà di un vero e proprio “suicidio cellulare” che necessita di tante proteine dedicate che svolgono un complesso e delicato meccanismo a tappe con un preciso schema prestabilito e con consumo di energia. In condizioni normali, l’apoptosi avviene continuamente perché contribuisce al mantenimento del numero di cellule di un sistema; avviene ad esempio in un seno che si ritira dopo la fase di allattamento dove ci sono migliaia di cellule che si “sacrificano” perché sono di troppo o non più necessarie, lo stesso vale per l’utero dopo una gravidanza etc. Ovviamente la mancata regolazione dell’apoptosi può causare disordini da perdita di cellule, caratteristica di molte patologie neurodegenerative, oppure una crescita cellulare incontrollata che è alla base delle neoplasie (tumori).
L’apoptosi, a differenza della necrosi, non causa infiammazione e la morte delle cellule mediante questo processo provoca la formazione di vescicole con un contenuto prestabilito di sostanze che possono essere riciclate da cellule specializzate. Dato che non c’è la rottura della membrana cellulare e che tutte le cellule del nostro organismo conoscono questo processo, l’apoptosi è un processo silenzioso che non fa danno.
L’infarto cerebrale: La scena del crimine
 Seguendo la storia sulla falsa riga del “giallo”, la prima domanda è come sono morte le cellule? Hanno eseguito un suicidio di massa (apoptosi) oppure sono morte per trauma (necrosi)?
Seguendo la storia sulla falsa riga del “giallo”, la prima domanda è come sono morte le cellule? Hanno eseguito un suicidio di massa (apoptosi) oppure sono morte per trauma (necrosi)?
I ricercatori con un po’ di esperienza sanno bene che un fenomeno di morte in massa è attribuibile solamente alla necrosi, l’apoptosi, infatti, è un fenomeno prevalentemente sporadico, ciò vuol dire che l’apoptosi in natura colpisce poche cellule alla volta e la loro scomparsa è quasi sempre impercettibile, mentre la necrosi è una morte cellulare a cascata, ogni cellula muore poiché si trova vicino una cellula in decomposizione ed è tipico della gangrena, dove si deve amputare un arto per frenare la morte a cascata che potrebbe propagarsi sul corpo del paziente e causarne la morte.
Possiamo essere sicuri che si tratti di necrosi?
Non esistono saggi specifici per la necrosi, ma per esserne sicuri potremmo usare sul cervello ischemico un anticorpo che riconosca selettivamente dei cartelli che indicano “qui c’è materiale apoptotico riciclabile” (epitopo). Se lo facessimo il risultato sarebbe deludente, il segnale apoptotico è molto scarso, l’apoptosi è avvenuta solamente su una piccola percentuale di cellule. E poi basta guardare come si presenta il materiale ischemico; si tratta di una zona amorfa in cui non è possibile riconoscere strutture macroscopiche e microscopiche, tranne alcune cellule resistenti che sopravvivono, e questo ci indica inevitabilmente che si tratta di una massa in necrosi… omicidio quindi.
Se si fosse trattato di una apoptosi di massa?
Se si fosse trattato di apoptosi (suicidio) ci saremmo aspettati qualche cellula qua e là morta con le classiche vescicolette, epitopo dell’apoptosi, assenza di infiammazione e sarebbe stato difficile identificarlo poiché silente.
Le cellule del nostro cervello
 L’investigazione quindi è tutta in salita, non possiamo trarre molte informazioni dal ritrovamento delle vittime se non che siano state uccise da un evento molto traumatico avvenuto diverse ore dopo il ristabilirsi delle condizioni ottimali di circolazione sanguigna (vedi post precedente).
L’investigazione quindi è tutta in salita, non possiamo trarre molte informazioni dal ritrovamento delle vittime se non che siano state uccise da un evento molto traumatico avvenuto diverse ore dopo il ristabilirsi delle condizioni ottimali di circolazione sanguigna (vedi post precedente).
Passiamo quindi alla raccolta delle informazioni sulle vittime nella loro vita quotidiana per poi passare alle indagini vere e proprie sull’assassino. Le vittime di questa storia sono ovviamente le cellule che partecipano alla struttura e funzione del cervello e non stiamo parlando solamente di neuroni. Il cervello, infatti, è costituito da centinaia di tipologie cellulari con ruoli distinti e diversamente collegati a tutte le altre cellule circostanti. Nel cervello abbiamo i neuroni che generano i nostri pensieri, che stimolano i muscoli a contrarsi e che generano anche ormoni che modificano il nostro stato fisico e psitico; poi abbiamo le cellule gliali, una famiglia di cellule nutrici per i neuroni che coccolano, proteggono, danno da mangiare e sostengono le cellule neuronali nel loro faticoso ruolo.
Le cellule gliali: Profilo di una serie di badanti ideali
Le cellule gliali sono suddivise in centinaia di ruoli distinti, dai più elementari ai più complessi, e sono anche le uniche cellule del cervello in grado di riprodursi, contrariamente ai neuroni. Proprio questa caratteristica è la prima distinzione tra le due popolazioni, mentre le cellule gliali possono moltiplicarsi e quindi invadere le zone morte in seguito ad un trauma, i neuroni possono solo diminuire il loro numero durante la nostra vita e mai aumentare. Pensate che il numero di cellule della glia in un cervello umano normale supera di nove volte quello dei neuroni, e proprio da questo è nato il falso mito che noi utilizziamo solamente il 10% del nostro cervello per pensare.
Alcune cellule della glia agiscono principalmente da supporto ai neuroni, altre regolano l’ambiente interno del cervello in particolare i fluidi che circondano i neuroni e le loro sinapsi provvedendo quindi al nutrimento delle cellule nervose. Alcuni tipi di cellule della glia producono molecole in grado di influenzare la crescita degli assoni. Altre funzioni sono state scoperte solo recentemente come la possibilità di comunicare con altre cellule.
La principale suddivisione della glia del cervello prevede due categorie principali:
Microglia: cellule specializzate nel mangiare e riciclare i rifiuti, come le cellule apoptotiche e gli agenti che disturbano la quiete del cervello. In genere si spostano all’interno del nostro cervello per rimuovere i corpi apoptotici e per controllare che tutto funzioni bene. In condizioni fisiologiche la microglia è presente solo in piccolo numero ma si moltiplicano in caso di danni neuronali.
Macroglia: una serie di cellule diverse, tra cui astrociti, oligodendrociti, cellule ependimali, glia radiale. I ruoli sono i più diversi tra loro e tutti hanno il comune denominatore di aiutare i neuroni a vivere in un ambiente ideale.
Bhé sicuramente se stiamo cercando un assassino non si troverà tra le vittime, tantomeno nella glia deputata a coccolare e proteggere i neuroni.
Neuroni: profilo di una vittima ideale

Anche i neuroni sono suddivisi in centinaia di sottotipi cellulari, ognuna con funzioni diverse e specifiche, ma tutti i neuroni hanno caratteristiche molto simili tra loro, tanto che possiamo accomunarli tutti in una sola descrizione generale.
I neuroni sono delle vittime ideali, poiché a differenza di tutte le cellule delle nostro organismo non si replicano mai, necessitano di molte cure e basta poco per ucciderle. Tutte le cellule del nostro organismo, infatti, possiedono delle riserve energetiche che sono utili in caso di prolungata assenza di nutrimenti, alcune cellule possono anche adattarsi a metabolizzare sostanze di diversa natura per sopravvivere ed altre ancora si adattano anche alla scarsa presenza di ossigeno. Purtroppo i neuroni fanno eccezione, non hanno riserve energetiche e si nutrono solamente di zucchero raffinato. Quel che è peggio è che non sanno adattarsi diversamente e che basta anche una carenza di ossigeno, sangue o alcune stimolazioni per innescarne la morte.
La vita di un neurone, inoltre, non è per niente facile, la sua membrana plasmatica è simile ad una batteria con una differenza di potenziale, espressa in milliVolt, che cambia in funzione dello stato cellulare, canali che fanno entrare ed uscire ioni modificando continuamente lo stato d’ordine cellulare mentre una serie di pompe ioniche cercano continuamente di ripristinare l’ordine con una certa fatica.
Tutti i neuroni sono poi interconnessi tra loro con una fitta rete di cavi elettrici (assoni e dendriti) che propagano o captano variazioni delle differenze di potenziale di membrana che viaggiano lungo un neurone, tra un neurone e l’altro e da un lato del cervello ad un altro.
Questi processi sono continui anche durante il sonno, durante la vita intrauterina e nel coma.
Se ne deduce che ognuno dei circa 100 miliardi di neuroni che abbiamo alla nascita è continuamente sull’orlo di un baratro con attività che lo spingono a fare cose eccezionali in frazioni di secondo, e tantissime proteine che devono continuamente ripristinare lo stato d’ordine in altrettanti frazioni di secondo con un poderoso dispendio di energia. Pensate che circa il 20% dell’energia che mangiamo, e che serve per farci stare in uno stato “normale”, serve semplicemente per far funzionare il cervello che rappresenta solo il 2% del peso corporeo; un’altra buona quota serve per mantenere ordine nel resto dell’organismo 98% del peso corporeo e solo una frazione piccola è necessaria per l’attività fisica che facciamo.
Conclusioni
In questo lunghissimo post abbiamo introdotto meglio alcuni concetti fondamentali che ci serviranno nei prossimi capitoli dove, se vorrete, cercheremo di prendere in considerazione alcune teorie su cosa sia avvenuto negli ultimi istanti di vita delle vittime, chi era presente al momento della morte, al momento del ritrovamento e soprattutto chi sia stato l’esecutore materiale di questo misterioso delitto, da dove proviene e come potremmo impedire che questo accada.
Tags:
Apoptosi,
Calcio,
Glia,
Ictus,
Ischemia Cerebrale,
Necrosi,
Neuroni
10 agosto 2008 - 8:15 pm
Rieccoci qua dopo un po’ di tempo di “meditazione lavorativa”, spero di recuperare i post di Nico, poiché sto un po’ indietro.
Allora, per ricominciare vorrei trattare un argomento che ha anche un importante risvolto filosofico che spero possa interessare anche ai “non addetti ai lavori”. Dovete sapere che quando ho iniziato a scrivere l’argomento qui di seguito ho scritto talmente tanto che mi sono subito reso conto che potrebbe essere un po’ troppo lungo e barboso per essere digerito in una sola volta. Di conseguenza ho deciso di farne un argomento a brevi “puntate”, spero che questo sistema vi sarà gradito maggiormente.
Se vi sono commenti vi prego di farmene partecipe.
Buona lettura
Pasquale

Per introdurre questo lungo argomento definirò prima il termine “ischemia“. La parola deriva dal Greco ισχαιμία, ovvero isch (riduzione) ed haima (sangue) e sottintende un apporto di sangue insufficiente per soddisfare le richieste energetiche di un determinato distretto dell’organismo. Ovviamente, la mancanza di nutrienti e di ossigeno porta prima alla sofferenza cellulare e poi, se è prolungata, determina la morte dei tessuti circostanti. L’estensione, inteso come volume, e l’entità del danno, inteso come percentuale di cellule morte, dipendono dal tipo cellulare coinvolto e dalla differenza tra la richiesta energetica e l’energia effettivamente fornita.
Il termine ischemia è riferibile a qualsiasi zona dell’organismo; per cui si può parlare di ischemia cardiaca, renale, muscolare, cerebrale etc. Ognuno di questi organi ha caratteristiche diverse, e di conseguenza sono più o meno sensibili al danno ischemico.
Con il termine ictus (dal latino “colpo”) si intende specificamente l’ischemia che riguarda il cervello. Questa patologia è la prima causa di invalidità permanente e la terza causa di morte nei Paesi industrializzati. I recenti progressi medico-scientifici hanno portato a dei miglioramenti nella gestione dei pazienti ischemici come la diagnosi precoce, la trombolisi, la creazione di centri per lo stroke e la riabilitazione. Tuttavia, l’ictus oggi rappresenta ancora un problema medico senza un’efficace terapia. Stranamente, nonostante i numerosi studi sull’argomento in questione l’ischemia cerebrale è ancora una patologia con molti lati oscuri da chiarire.
L’ictus, infatti, presenta diverse caratteristiche anomale che lo contraddistinguano dagli altri tipi di ischemia. La caratteristica più evidente è che la morte dei neuroni non avviene quando c’è la carenza di carburante ed ossigeno, ma diverse ore fino a giorni dopo il ristabilirsi del normale flusso sanguigno. Si può affermare quindi che la morte neuronale non è causata direttamente della semplice mancanza di cibo ed ossigeno, ma per complessi meccanismi che sono scaturiti da questa mancanza. Questa caratteristica rende l’ischemia cerebrale una sezione medica di notevole interesse scientifico poiché è una patologia molto grave e diffusa, con un periodo di possibile intervento (finestra terapeutica) abbastanza ampia. Il tempo che intercorre tra l’ictus ed il raggiungimento del massimo danno cerebrale è abbastanza lungo per un tempestivo intervento farmacologico sul paziente. Purtroppo le attuali terapie mirano a fenomeni macroscopici come ripristinare il flusso sanguigno mediante agenti anticoagulanti che rompono il coagulo che ha occluso un’arteria. Queste terapie sono utili solo se sono effettuate entro 2-3 ore dall’inizio dei sintomi ed hanno scarsa efficacia. Secondo molti la “vera terapia” dovrebbe mirare a bloccare quei sistemi cellulari che portano una determinata cellula, apparentemente non sofferente, a morire dopo giorni dall’evento ischemico. Per questo motivo è in corso da tempo la caratterizzazione di questi complicati meccanismi cellulari per la ricerca di efficaci bersagli farmacologici che siano in grado fermare questo percorso neurodegenerativo ritardato. Sin dai primi risultati delle ricerche svolte è emerso che uno dei punti chiave della neurodegenearazione è lo ione calcio e la fine regolazione delle sue concentrazioni intracellulari ed extracellulari.
Il Calcio: la “Spada di Damocle” dell’ischemia

Il Calcio (Ca2+) è un componente essenziale per la vita; questo ione infatti oltre ad essere un componente fondamentale per le ossa, è usato anche come un importante segnale cellulare per innescare migliaia di processi fisiologici che vanno dalla contrazione cellulare, come nel caso delle cellule cardiache e muscolari, fino all’impulso nervoso di ogni singolo neurone del cervello.
Pensate che la concentrazione di calcio presente all’interno di una qualsiasi cellula del nostro organismo a riposo è estremamente bassa (50-100 nanoMolare), praticamente meno della concentrazione di calcio che si ha nella semplice acqua distillata! All’esterno delle cellule, invece, c’è una concentrazione di calcio di circa 10.000 volte superiore che è separata dall’interno delle cellule da un doppio strato lipidico sottilissimo. Questa differenza di concentrazione è fondamentale, poiché basta anche un lieve incremento dei livelli di calcio intracellulari per innescare in base al tipo di cellule interessate: l’apertura di canali, fusioni di vescicole, contrazioni cellulari, depolarizzazioni nervose, rilascio di neurotrasmettitori e/o ormoni. Una cellula neuronale, durante la sia attività, può passare per brevi istanti da concentrazioni di calcio di 50-100 nanoMolare fino a 1 milliMolare (1.000 volte di più) per poi ritornare ai livelli basali e prepararsi per essere stimolata nuovamente.
Potete immaginare che mantenere bassa la concentrazione di calcio all’interno delle cellule richiede molti sforzi energetici che consumano ATP, la molecola che conserva l’energia cellulare. Immaginate ora cosa può accadere quando manca il sangue in un determinato distretto del cervello; la mancanza di ossigeno e nutrimenti non consentirebbero alle cellule di produrre energia sottoforma di ATP e di conseguenza comincerebbero ad essere meno efficienti tutti i meccanismi che servono a mantenere basse le concentrazioni cellulari di Calcio. In definitiva, durante l’ischemia, i neuroni non riescono più a cacciare fuori il calcio che entra durante la propria attività, di conseguenza anche le funzioni fondamentali sono compromesse ed inizia la lunga agonia che porterà alla morte cellulare.
Il Calcio, quindi, è una spada di Damocle che pende su tutte le cellule che ne fanno uso; l’ingresso di calcio è fondamentale per attivare la maggior parte dei meccanismi cellulari, ma quando un neurone non c’è l’energia necessaria per estruderlo per ritornare nuovamente alle condizioni basali, le concentrazioni citoplasmatiche di questo ione aumentano e di conseguenza si innescano, in maniera incontrollabile, una serie di meccanismi non desiderati con un ulteriore dispendio energetico.
A complicare le cose c’è la fitta rete di contatti che i neuroni hanno tra di essi e con l’organismo. L’accumulo di calcio in un neurone provoca un’attivazione incontrollabile con il rilascio di neurotrasmettitori eccitatori verso altri neuroni, che a loro volta si attiveranno facendo entrare ioni calcio che non riusciranno ad estrudere. La reazione a catena potrebbe continuare all’infinito in tutto il cervello causando la generazione di scariche elettriche senza controllo che si propagano longitudinalmente rispetto il centro dell’infarto cerebrale (core ischemico) con liberazione di neurotrasmettitori e conseguenze che vedremo nelle prossime puntate.
Il calcio quindi è uno ione fondamentale per la vita perché è il centro di regolazione di funzioni cellulari importanti. Tuttavia proprio per questo motivo è anche uno ione molto pericoloso perché in condizioni patologiche, come l’ictus per l’appunto, può innescare l’autodistruzione cellulare. Un articolo pubblicato su Journal of Neuroscience descrive l’importanza di alcuni meccanismi; per chi è interessato
Durukan and Tatlisumak Pharmacology, Biochemistry and Behavior 87 (2007) 179-197.
Tags:
Biologia molecolare,
Ictus,
Ischemia Cerebrale,
Mente,
Neuroscienze,
Neurotrasmettitori
7 gennaio 2008 - 3:20 pm
Uno dei motivi per cui leggo spesso le riviste con altissimo impact factor è la speranza di trovare un articolo davvero rivoluzionario che possa chiarire un determiato meccanismo oppure possa aprire la strada verso nuove prospettive, oltre che per aggiornarmi sulle recenti scoperte. Purtroppo devo dire che le riviste scientifiche più importanti alla fine non pubblicano sempre cose drasticamente innovative o chiarificatrici, anzi a volte le riviste un po’ più modeste contengono articoli con informazioni molto più convincenti e serie rispetto a Nature, Cell, Neuron etc.
In una di queste letture, mi è capitato di leggere qualcosa su Cell che, a mio avviso, è veramente incredibile, oltre che curioso. L’articolo in questione tratta di un canale del calcio voltaggio dipendente (Cav1.2) che ha anche un secondo ruolo funzionale del tutto diverso ed imprevedibile da quello che ci si aspetterebbe da un semplice canale.
Prima di entrare nel merito bisognerebbe introdurre il concetto di canale voltaggio dipendente oramai acquisito da tanti anni nel campo delle neuroscienze ma che per i non addetti ai lavori potrebbe sembrare fuorviante.
Diciamo che almeno qualche migliaio dei circa 30.000 geni umani codificano per proteine di membrana che possono formare dei canali idrofilici attraverso cui, in particolari condizioni, possono passare degli ioni. Il motivo per cui esistono tante proteine non è dovuto alla presenza tanti tipi di ioni da far passare, ma dalla fine regolazione a cui questi canali devono rispondere. La quasi totalità di questi canali ionici sono sempre chiusi, ma possono aprirsi in determinate condizioni secondo una cinetica caratteristica che è dipendente dai componenti da cui è formato.
Alcuni canali possono aprirsi solamente in seguito al legame di un neurotrasmettitore sulla regione esterna, o interna, del complesso proteico (Recettori canale), come avvine per il recettore nicotinico che fa entrare sodio all’interno della cellula in seguito al legame dell’acetilcolina con la parte esterna del recettore; la cinetica di apertura/chiusura o l’affinità con l’agonista recettoriale dipende dai componenti del canale stesso. Ovviamente l’ingresso di sodio che ne scaturisce provoca a sua volta l’attivazione di una serie di altri meccanismi a cascata come le tessere del domino che portano all’attivazione di altri effettori. Altri canali possono invece aprirsi in seguito al cambiamento delle condizioni cellulari, variazione di pH (es ASICs), di concentrazioni ioniche (es Connessine) oppure del potenziale di membrana (es canali voltaggio dipendenti).
Ogni tipo cellulare possiede il proprio corredo di recettori canale, canali voltaggio dipendenti, canali passivi e trasportatori; l’insieme di tali complessi proteici con caratteristiche finemente diverse tra loro determina delle differenze macroscopiche e microscopiche nella reattività cellulare, forma del potenziale d’azione, velocità di risposta ad uno stimolo, eccitabilità di membrana, potenziale di membrana a riposo, picco di depolarizzazione e tantissime altre variabili che fanno di un neurone un sottotipo cellulare unico. Non entro nel dettaglio di quanti canali del calcio voltaggio dipendenti esistono, dico solo che sono identificate in varie classi con la sigla Cav seguita da un numero che identifica la classe seguito da un punto e poi un altro numero che identifica il sottotipo. Tutti questi canali regolano direttamente o indirettamente tante funzioni neuronali, come la memoria, il dolore, l’eccitabilità, la fertilità e la sensibilità a determinati danni patologici. Tuttavia è noto anche che questi canali ionici possono avere anche una funzione nella regolazione della trascrizione genica, poiché molti fattori trascrizionali sono regolati dalle concentrazioni di determinati ioni come ad esempio il calcio. Questa è stata sempre una teoria debole, poiché indubbiamente ci sono dei casi in cui l’effetto sulla trascrizione genica può essere drasticamente diverso se si trasfettano in cellule dei canali composti da subunità diverse. Insomma se è l’ingresso di ioni calcio a determinare l’attivazione di alcuni fattori trascrizionali allora l’effetto dovrebbe essere uguale per qualsiasi canale che fa entrare calcio, soprattutto se 4 subunità su 5 sono identiche e le caratteristiche cinetiche di ingresso e uscita del calcio sono praticamente le stesse.
Tuttavia questo non avviene sempre, come ho già accennato, l’effetto può essere drasticamente diverso, tale che alcuni canali ionici possono determinare il differenziamento neuronale in alcune cellule, mentre altri canali possono determinare l’espressione di proteine del metabolismo o neurotrasmettitori in maniera non chiara.
Una spiegazione che, secondo me, ha aperto la strada e gli occhi a molti ricercatori scientifici risiede nell’articolo di cui parlavo all’inizio di questa disquisizione, pubblicato su un numero di Cell dell’anno scorso dalla dott.sa Gomez-Ospina dell’università di Stenford. Il canale del calcio voltaggio dipendente (Cav1.2) possiede nel dominio C-terminale una sequenza peptidica che, se tagliata, è di fatto un fattore trascrizionale a tutti gli effetti e modula un cospicuo gruppo di geni endogeni importanti per la neurotrasmissione e per l’eccitabilità neuronale.
Il fattore trascrizionale in questione è stato definito Fattore Trascrizionale Associato al Canale del Calcio (CCAT) e si origina mediante la proteolisi di Cav1.2. Appena liberato, questo fattore trascrizionale si localizza nel nucleo dei soli neuroni grazie a meccanismi non ancora ben chiari e lì lega proteine nucleari e sequenze specifiche di DNA attivando la trascrizione dei geni target.
La cosa curiosa è che CCAT a sua volta è regolato dalle concentrazioni di calcio e dalla presenza di altri fattori trascrizionali che variano durante lo sviluppo, quindi il suo funzionamento è regolato da diverse variabili. L’effetto finale che ha il canale Cav1.2 quindi non è semplicemente variare la concentrazione di calcio ma regolare direttamente la trascrizione di geni target.
Bhé in definitiva questo articolo ci racconta come a volte le teorie che vengono formulate e che leggiamo di continuo non siano altro che un velo oltre il quale si nascondono dei meccanismi, come in questo caso, inimmaginabili e forse anche al limite della credibilità secondo le attuali conoscenze molecolari.
Forse non tutti i meccanismi sono di questo tipo e così semplici da spiegare, ma possiamo essere certi che c’è ancora tanto da scoprire dietro le nostre attuali teorie.
Tags:
Biologia molecolare,
Canali voltaggio dipendenti,
fattori trascrizionali,
Neuroscienze,
Potenziali d'azione
 Commenti disabilitati
Commenti disabilitati
21 dicembre 2007 - 11:59 pm
 Nel 1983 il genio Woody Allen porta sul grande schermo una storia strana che diventerà un cult. Trattasi del film comico Zelig che ha poi ispirato il famoso omonimo spettacolo italiano dei comici del venerdì sera.
Nel 1983 il genio Woody Allen porta sul grande schermo una storia strana che diventerà un cult. Trattasi del film comico Zelig che ha poi ispirato il famoso omonimo spettacolo italiano dei comici del venerdì sera.
Per chi non avesse mai visto o sentito parlare di questo film, si può definire come un “documentario” comico sulla vita di uno strano individuo, di nome Leonard Zelig (interpretato da Woody Allen), che privo di una propria memoria cerca inconsapevolmente di acquisire l’identità di chi gli sta intorno. Una storia ambientata nel 1920, quindi in bianco e nero, che racconta tra l’altro anche una piccola storia d’amore tra il protagonista e la dottoressa che lo tiene in cura (interpretata da Mia Farrow). La scrittura di questa storia si basò sulla pura fantasia di Woody Allen, ma oggi a distanza di tanti anni si scontra all’improvviso con la realtà di uno strano caso clinico neurologico.
Nella psichiatria, infatti, esistono numerosi casi “strani”, e non sto parlando del classico tipo che si identifica in Napoleone, Cleopatra oppure uno delle migliaia di personaggi più o meno noti; questi casi, infatti, presentano una identificazione univica di un personaggio rifiutando la propria identità personale per un tempo prolungato. Nell’ambito della patologia psichiatrica c’è anche un’altra classificazione che va sotto il nome di dipendenza ambientale, anche detta sindrome d’uso o comportamento di utilizzazione, in cui i pazienti imitano i gesti dei loro interlocutori o tendono ad usare tutti gli oggetti che hanno davanti.
Sulla rivista inglese Neurocase, tre noti psichiatri napoletani (Giovannina Conchiglia, Gennaro Della Rocca e Dario Grossi) hanno descritto un caso, che hanno avuto recentemente tra le loro mani, unico nel suo genere. Il loro paziente è malato di un disturbo comportamentale che è stato definito Zelig-like syndrome. Quello che distingue questo paziente dai casi precedentemente riportati è la sua totale e onnipervasiva immersione in un contesto con un adattamento eccessivo a ruoli, e non a semplici stimoli, proposti di volta in volta dall’ambiente che lo circonda. Il tutto si manifesta con la perdita della capacità di mantenere costante la propria identità.
Il paziente in questione è un ex-professionista napoletano di 65 anni (con cui mi riferirò con il nome di fantasia Leonard) che in seguito ad un arresto cardiaco, che gli ha provocato un’ipossia cerebrale con danni al lobo fronto-temporale, ha dei disturbi comportamentali che lo fanno assomigliare molto al personaggio ideato da Woody Allen.
Ovviamente si tratta di un caso senza precedenti ed unico nel suo genere; Leonard riesce inconsapevolmente a trasformarsi nel suo interlocutore diventando medico quando interagisce con un medico, esperto di cocktail quando si trova di fronte ad un barman ed esperto di culinaria quando è in contatto con dei cuochi. Ovviamente non si tratta di un semplice caso di pazzia come si potrebbe dedurre da queste semplici frasi, l’immersione di Leonard nel contesto in cui di volta in volta si trova è pressoché totale. Gli psicologi hanno eseguito una serie di test dai risultati sorprendenti. Quando era barman, a chi gli chiedeva come si preparasse un determinato cocktail, ha risposto di essere ancora in prova: «Sono qui da due settimane, spero di avere il posto fisso». In cucina era un cuoco provetto: «Sono uno chef specializzato in menu per diabetici», ha spiegato senza un’ombra di esitazione, assolutamente immedesimato nella sua nuova identità. L’unico ruolo in cui non si è calato è stato, chissà perché, quello di addetto alla lavanderia della casa di cura. Ma per il resto, Leonard ha “rubato” il mestiere a tutte le persone che aveva davanti.
Una sorta di trasformismo psicologico coatto, di camaleontismo involontario che lo rende di volta in volta un perfetto giocatore di baseball in mezzo ai campioni di baseball, un trombettista nero in una banda di jazz, un pellerossa tra i pellerossa, un ebreo tra gli ebrei, uno psicanalista fra gli psicanalisti e cardiologo fra i cardiologi. In ogni occasione Leonard cerca anche di usare un linguaggio appropriato al ruolo che riveste. La dott.ssa Conchiglia ha descritto le domande “trabocchetto” a cui ha sottoposto il paziente, per esempio al cardiologo che gli ha chiesto a quale patologia corrispondesse una determinata anomalia del battito cardiaco, Leonard ha replicato in modo generico ed evasivo ma quanto più appropriato possibile, del tipo “La domanda è troppo complessa, dipende da paziente a paziente”. Da perfetto Zelig, Leonard usa inconsciamente una formidabile arma di difesa contro il suo involontario trasformismo: cancella totalmente dalla memoria il ruolo che ha appena sostenuto quando si immedesima in uno nuovo. Quando è un medico non è mai stato un barman o un libero professionista, nè sa dire nulla di quando sosteneva di saper cucinare alla perfezione.
La dott.ssa Conchiglia ci tiene a precisare anche che, durante i numerosi e repentini cambi di identità, l’uomo-camaleonte, non perde mai il suo carattere e la sua personalità; quel che maggiormente colpisce, piuttosto, è la capacità di adattarsi ai contesti sociali più diversi in cui viene a trovarsi. C’è da dire anche che il paziente non ha mai una completa amnesia, Leonard riconosce e ricorda quasi sempre la moglie ed i figli che non lo perdono di vista per un solo istante.
Probabilmente non guarirà mai, anche se le terapie in day hospital a cui è sottoposto hanno consentito un lieve miglioramento delle sue condizioni. Anche se le crisi sono meno frequenti, l’uomo-camaleonte non è certo in grado condurre un’esistenza normale, e di conseguenza anche la sua autonomia è limitata.
Questo caso napoletano ha avuto un certo eco nel mondo delle neuroscienze e non solo, recentemente nella serie americana Dr. House, portata anche in Italia su canale 5, uno degli autori ha preso ispirazione da questo caso per una delle nuove puntate (titolo inglese “Mirror mirror”) che arriverà prossimamente anche in Italia nella quarta serie. Il burbero medico diagnostico si troverà infatti di fronte ad un paziente con una sindrome clinica simile a quella recentemente identificata (la Giovannini’s syndrome). Lascio a voi vedere come andrà a finire.
Tags:
Casi clinici,
Memoria,
Mente,
Neuroscienze,
Psicologia
