9 febbraio 2009 - 9:19 pm
Ciao a tutti! Oggi vi parlerò di un articolo che non tratta strettamente le neuroscienze, ma che propone una tecnica che sicuramente potrà trovare interessanti risvolti in questo campo. Questo post è la traduzione (abbastanza fedele) di quello che ho scritto originariamente (in inglese) sull’ottimo blog ReporterGene.
Il
calcium imaging è una tecnica che è decisamente maturata negli ultimi tempi, e di anno in anno vengono generati indicatori per il
calcio codificati geneticamente sempre più evoluti.
Uno dei vantaggi dell’utilizzo di indicatori codificati geneticamente è quello di poterli utilizzare per l’imaging in vivo e quindi per studiare processi fisiologici (o patologici) in un sistema molto più completo (e complesso) di -per esempio- una coltura cellulare o una fettina di tessuto.
Diversi approcci sono stati utilizzati per ottenere imaging in vivo, alcuni dei quali fanno uso di una fibra ottica, o “fibroscopio”, inserita nella regione da analizzare e collegata ad un microscopio. Questo è ovviamente un metodo invasivo, e limita l’osservazione alla piccola zona d’azione della fibra. Un altro approccio meno invasivo è quello di utilizzare obiettivi a lunga distanza focale per ottenere immagini di una regione del corpo esposta: come già visto, questo metodo è stato usato, ad esempio, per analizzare la plasticità delle spine dendritiche nella corteccia cerebrale. Anche questa tecnica, tuttavia, permette la visione di una limitata area di interesse, oltre a richiedere un intervento chirurgico piuttosto delicato.
L’approccio recentemente descritto da Rogers e colleghi su PlOS One, invece, è completamente non invasivo. Hanno infatti generato una linea di topi transgenici che esprimono in maniera selettiva (tamite un approccio floxed-stop) una proteina sensibile al calcio, detta GFP-aequorina. Il reporter può essere espresso selettivamente nelle cellule di interesse semplicemente incrociando questi topi con topi che esprimono Cre in quelle cellule (molte di queste linee sono disponibili commercialmente). La GFP-aequorina è una proteina che consiste di una parte sensibile al calcio (aequorina) che in presenza di ioni Ca2+  può ossidare un substrato (chiamato coelenterazina), producendo luminescenza, la quale eccita l’altra metà della proteina (GFP, green fluorescent protein) che quindi fluoresce.
può ossidare un substrato (chiamato coelenterazina), producendo luminescenza, la quale eccita l’altra metà della proteina (GFP, green fluorescent protein) che quindi fluoresce.
L’idea è quindi quella di iniettare coelenterazina e misurare la fluorescenza generata da GFP.
Questo metodo permette di osservare variazioni di calcio nell’intero animale, anche se ovviamente ha una risoluzione spazio-temporale più bassa (ma comunque buona) rispetto agli altri metodi sopra menzionati.
E in effetti sembra che funzioni bene! Quando GFP-aequorin è stata espressa nei mitocondri delle cellule muscolari, infatti, ha permesso di misurare l’aumento di calcio durante la contrazione muscolare. Questo è stato testato sia dopo una contrazione introdotta artificialmente, sia in condizioni fisiologiche come l’attività muscolare spontanea di topi neonati (vedi figura), sia in situazioni “patologiche”, come le convulsioni evocate da kainato.
Per meglio vedere questo sistema in azione consiglio fortemente di guardare i video allegati all’articolo (che come tutti gli articoli di PLoS è ad accesso gratuito).
Riassumendo, questo approccio permette di misurare il calcio in modo non invasivo e su tutto l’animale in condizioni fisiologiche e patologiche. Questa tecnica potrebbe trovare interessanti applicazioni in studi che misurano lo stato metabolico di un certo tessuto, o l’apoptosi o uno dei moltissimi processi regolati dal calcio (la memoria ad esempio). Il passo successivo, ovviamente, sarebbe quello di ottenere questo tipo di registrazioni in un sistema in cui l’animale si può muovere liberamente. Se avete visitato il link qui a sinistra avrete capito che non siamo lontani dall’ottenerlo, ed infatti lo stesso gruppo ha già pubblicato un articolo a riguardo (Roncali et al., J. Biomed. Opt. 2008)!
Tags:
Animali transgenici,
Calcio,
Imaging,
In vivo
23 novembre 2008 - 11:53 pm
In post precedenti abbiamo già parlato di memoria. Oggi vi voglio parlare di un recente studio che ho trovato molto interessante, e che analizza come il sonno possa influenzare la memoria. Una delle teorie riguardanti il sonno dice che esso ci permette di consolidare gli avvenimenti del giorno e formare nuove memorie grazie al riarrangiamento delle sinapsi fra i neuroni di diverse zone cerebrali. Un recente articolo di Susanne Diekelmann e colleghi, pubblicato su PlOS One, correla la deprivazione di sonno con la formazione delle cosiddette false memorie. Trovate il testo completo (accessibile gratuitamente) qui: “Sleep Loss Produces False Memories“.
Innanzitutto definiamo cosa si intende per falsa memoria: si intende il ricordo da parte di una persona di un fatto falso, di qualcosa che non è mai avvenuto, senza però che il soggetto stia mentendo; anzi, la persona è assolutamente in buona fede in quanto -almeno nella sua memoria- quello specifico avvenimento c’è effettivamente stato. L’ovvia domanda a questo punto è: come è possibile che vengano a formarsi queste false memorie?
Il processo di formazione della memoria ci è ancora in parte oscuro, ma sembra che il sonno sia una componente essenziale nel plasmarla. Durante il giorno noi assistiamo a degli avvenimenti, impariamo nuove nozioni, facciamo nuove scoperte e di notte il nostro cervello processa il tutto, escludendo alcune cose, e salvandone altre. Questo processo consiste di una consolidazione sinaptica, per cui i nuovi fatti che abbiamo imparato vengono stampati in particolari circuiti cerebrali, e di una consolidazione del sistema, per cui questi nuovi fatti vengono integrati con fatti preesistenti e già memorizzati in altri circuiti. Questo ultimo processo ci permette di correlare le nostre vecchie memorie con quelle nuove.
Una delle ipotesi sulle false memorie è quella che, poichè il nostro cervello non è una macchina perfetta, lo stesso processo di memorizzazione possa portare alla formazione di false memorie, se il consolidamento di questi nuovi dati non viene effettuato correttamente. Lo studio di cui voglio parlarvi oggi, tuttavia, analizza la situazione opposta, cioè la deprivazione del sonno, come fattore che può generare false memorie.
Lo studio in questione ha comparato la formazione di false memorie in diversi gruppi di persone deprivate o meno del sonno in diverse situazioni.
Ma come si può valutare una cosa del genere? Esiste un apposito test, chiamato test DRM (dalle iniziali dei suoi inventori: Deese, Roediger e McDermott) che massimizza la formazione di queste false memorie ed è quindi un buon metodo per valutarne la presenza. Il test è molto semplice: nella prima fase di training vengono lette ai partecipanti vari gruppi di 15 parole in qualche modo correlate, ad esempio: “notte”, “buio”, “carbone”, … In questa lista, tuttavia, manca il termine chiave, nel nostro esempio “nero”, che li collega tutti. Dopo un certo periodo ai partecipanti vengono mostrate delle parole rientranti in 3 categorie: le parole della lista, che erano presenti tra quelle lette nella fase di training, parole trabocchetto, cioè le parole chiave (“nero” nell’esempio di prima) che sono collegate alle altre ma non erano state lette, ed infine parole di altre liste, ad es. “alto”. I partecipanti devono quindi dire se le parole mostrate erano state lette loro in precedenza, e dare anche un’indicazione (come punteggio da 1 a 4) su quanto siano sicuri della propria risposta. Ovviamente, poichè le parole chiave sono correlate alle altre, è facile che una persona ne abbia una falsa memoria. Il test dà quindi tre risultati: false memorie (es. dire che “nero” era nella lista), hits, cioè risposte esatte (es. dire che “notte” era nella lista), e falsi allarmi (es. dire che “alto” era nella lista).
Per vedere se effettivamente il sonno ha un ruolo nella formazione di queste memorie, sono stati testati diversi paradigmi di deprivazione, mostrati in questa figura.
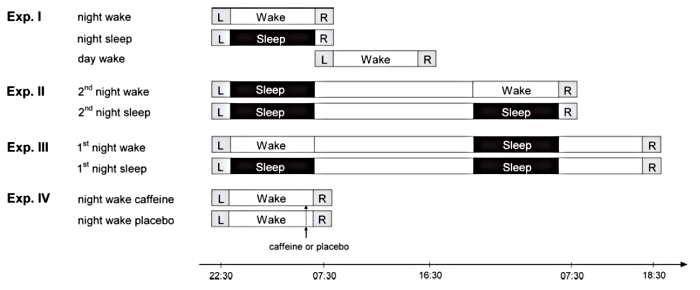
L’esperimento 1 consiste di 3 gruppi:
- un gruppo che riceve il training (indicato da L nell’immagine) di sera, rimane sveglio (wake) di notte e viene testato (quadrato con R) la mattina dopo.
- un gruppo che riceve il training sempre di sera, ma dorme (sleep) di notte, per essere poi testato al mattino.
- un terzo gruppo che riceve il training di mattina e viene testato il pomeriggio, senza aver dormito.
Il risultato di questo primo esperimento è molto interessante: il gruppo deprivato di sonno, infatti, mostra più false memorie non solo rispetto al gruppo che ha dormito, ma anche rispetto al gruppo testato durante il giorno. Non c’è invece differenza fra il gruppo che ha dormito e quello testato di giorno. I tre gruppi hanno invece mostrato percentuali non differenti di hits e falsi allarmi.
Questo sembrerebbe indicare che la deprivazione del sonno durante la notte provoca un aumento delle false memorie, ma che il sonno non sia strettamente necessario per consolidare le memorie, almeno a breve termine.
Con l’esperimento 2 il gruppo ha investigato più nel dettaglio questi processi.
In questo caso entrambi i gruppi ricevono il training di sera, dormono e passano la giornata successiva senza essere testati. A questo punto, un gruppo dorme e l’altro viene deprivato del sonno, prima del test la mattina successiva.
Anche in questo caso i risultati sono gli stessi dell’esperimento 1. Il gruppo che non ha dormito prima del test presenta una maggiore percentuale di false memorie (ma non di hits o falsi allarmi) rispetto al gruppo di controllo che ha dormito. Questo vuol dire che il problema sta nella fase di retrieval, cioè di recupero delle memorie al momento del test. Chi non ha dormito di notte fa più fatica a recuperare quello che il suo cervello ha memorizzato e quindi ha più false memorie.
Questi risultati sono stati confermati dall’esperimento 3, in cui la deprivazione del sonno viene effettuata in uno dei due gruppi dopo il training, ma non prima del testing. In questo caso non ci sono state differenze fra i due gruppi. Questo significa che anche chi non ha dormito dopo il training è stato in grado di consolidare le nuove memorie, ma avendo entrambi i gruppi dormito prima del test hanno avuto ugual capacità di recuperare queste memorie, confermando quindi l’importanza del sonno per il retrieval.
Infine, l’ultimo esperimento che è stato effettuato ha valutato il coinvolgimento di un particolare neurotrasmettitore, chiamato adenosina, che in altri studi è stato visto essere correlato a problemi cognitivi risultanti dalla deprivazione del sonno. L’azione dell’adenosina nel cervello può essere “controllata” somministrando caffeina, in quanto questa sostanza lega gli stessi recettori che vengono legati dall’adenosina (ovvero, la caffeina è un antagonista dei recettori dell’adenosina).
In questo esperimento tutti i soggetti hanno subito deprivazione del sonno ma, un’ora prima del test, a metà è stata somministrata caffeina ed all’altra metà un placebo. In questo caso il gruppo trattato con caffeina ha avuto performance simili a quelle di chi negli altri esperimenti aveva dormito, quindi con una percentuale di false memorie più bassa rispetto a quella di chi ha ricevuto il placebo. Questo indicherebbe una possibile influenza dell’adenosina nella formazione di false memorie in seguito a deprivazione del sonno. La quantità di caffeina somministrata in questo studio (una capsula da 200mg) può essere comparata più o meno a quella presente in due tazzine di espresso. Ovviamente, come è noto, la caffeina ha anche un effetto sull’attenzione; andrebbe quindi valutato se l’effetto è veramente dovuto all’attivazione del sistema adenosinergico o se altri stimolanti non specifici per i recettori dell’adenosina hanno gli stessi effetti.
Gli autori concludono quindi che la deprivazione del sonno porta ad un aumento delle false memorie, mentre il sonno dopo il training non ha effetto sulla loro formazione.
Sarebbe interessante a questo punto studiare l’effetto di lunghi periodi di deprivazione. Cosa succederebbe dopo 2 o 3 giorni che uno non dorme? E dopo una settimana? Ok, immagino che forse sarebbe complicato trovare i volontari per l’esperimento… ma sarebbe decisamente interessante!
Tags:
Adenosina,
Memoria,
Neurotrasmettitori,
Psicologia
11 novembre 2008 - 9:21 pm
Siete bravi in matematica? Se la risposta è sì, forse può dipendere dal fatto che abbiate una buona capacità matematica istintiva,
non formale. Insomma, che avete un buon “sesto senso” per approssimare le quantità. Questa la tesi proposta da Halberda e colleghi in un loro recente articolo, apparso su Nature.Potete trovare il testo originale qui (richiesto abbonamento alla rivista): “
Individual differences in non-verbal number acuity correlate with maths achievement”
 Lo studio ha comparato l’abilità matematica formale di 64 ragazzi di 14 anni con la loro abilità istintiva, ovvero il loro approximate number system (ANS) o sistema di conto approssimativo.
Lo studio ha comparato l’abilità matematica formale di 64 ragazzi di 14 anni con la loro abilità istintiva, ovvero il loro approximate number system (ANS) o sistema di conto approssimativo.
Per misurare questo indice i ricercatori hanno utilizzato un semplice test: ai ragazzi veniva presentata una schermata con un certo numero di pallini gialli e blu di diversa dimensione per un tempo di 200 ms, troppo breve per permettere un effettivo conteggio. A questo punto il soggetto doveva dire quali cerchi fossero più numerosi: quelli gialli o quelli blu. Ovviamente la disposizione e la quantità dei cerchi erano casuali, in modo da escludere un effetto della posizione. Un test molto semplice, che permette di valutare l’abilità istintiva a contare, che non necessita di alcuna abilità formale di matematica (es. non è necessario saper risolvere un’equazione per riuscire in questo test).
Come facilmente immaginabile, la percentuale di risposte giuste è correlata al rapporto fra il numero dei due gruppi: ad esempio, è più facile dare la risposta giusta avendo 10 cerchi blu e 5 gialli, che avendone 8 gialli e 9 blu. Questo fenomeno è noto come legge di Weber. La risposta dei singoli soggetti, tuttavia è stata estremamente variabile: alcuni soggetti sono stati in grado di misurare accuratamente quantità in rapporto 9:10 (es. 9 blu e 10 gialli), altri facevano fatica a discriminare rapporti più fini di 2:3 (es. 10 blu e 15 gialli).
La domanda che i ricercatori si sono posti è la seguente: può la capacità istintiva di contare essere la base di buoni risultati nella matematica formale? I ragazzi scelti per il test erano stati valutati nel corso degli anni scolastici precedenti per le loro capacità matematiche formali usando test matematici standardizzati. Il confronto fra i risultati di questi studi ha in effetti mostrato che la capacità matematica formale è correlata al livello di ANS. Il confronto con i risultati di altri 16 diversi test fatti sugli stessi soggetti, per valutare le loro abilità cognitive e comportamentali più “generali”, non hanno mostrato alcuna correlazione con il livello di ANS.
Gli autori concludono che poichè l’ANS è attivo durante l’infanzia, esso possa essere importante per lo sviluppo delle abilità cognitive. Alternativamente, la situazione potrebbe essere invertita: una maggiore quantità o qualità dell’insegnamento della matematica formale potrebbe infatti aumentare l’indice di ANS.
Questa ipotesi alternativa potrebbe spiegare differenze nell’acuità dell’ANS in popolazioni di adulti che hanno ricevuto un’educazione matematica migliore di altre.
Se volete testare la vostra capacità matematica informale, potete trovare un test a
questo indirizzo. Per avere un risultato “valido” ripetete il test almeno 25-30 volte. Io ho ottenuto un fantastico 89%! E voi?
Tags:
Matematica,
Mente,
Neuroscienze,
Psicologia
30 luglio 2008 - 5:54 pm
Molti dei nostri processi biologici sono regolati da ritmi regolari. Il nostro cuore batte circa 70 volte al minuto, molti ormoni vengono prodotti dal cervello ad intervalli regolari di 2-3 ore, e così via. Oltre a questi ritmi, spesso chiamati “ultradiani” (cioè che si ripetono più di una volta al giorno) esistono però dei ritmi “circadiani” cioè che si ripetono con cicli di un giorno, che regolano ad esempio il fatto che – normalmente – siamo attivi di giorno e dormiamo di notte. La luce gioca quindi un ruolo estremamente importante nel controllare queste funzioni.
Una delle aree che si ritengono essere più importanti per il controllo dei ritmi circadiani è il nucleo soprachiasmatico (SCN), una regione cerebrale che riceve afferenze anche dalla retina ed è quindi in grado di processare le informazioni sull’orario attuale (giorno/notte).
Oggi vi parlerò di un interessante studio pubblicato qualche anno fa su
Nature, nel quale si analizzava il ruolo di due geni importanti per il mantenimento di questi ritmi, chiamati cry1 e cry2.
La scoperta dei geni cry è già di per se molto interessante. Sono stati infatti scoperti come omologhi di geni che codificano per i criptocromi, pigmenti sensibili alla luce blu presenti in alcune piante e responsabili di processi quali la fioritura in determinati periodi dell’anno. Ma a cosa servono in un mammifero?
Lo studio di van der Horst e colleghi analizza il comportamento di topi knockout (KO) per questi geni, ossia topi a cui sono stati rimossi questi determinati geni. Quando confrontati con animali normali, infatti, i topi knockout per i geni cry presentano ovvie anomalie.

L’immagine qui sopra mostra l’attività di topi normali e KO per i geni cry. Le barrette nere indicano l’attività motoria dei topi, lo sfondo bianco indica luce e grigio indica buio. I topi normali (pannello a) sono animali notturni e come tali mostrano attività motoria di notte e non di giorno. L’attività motoria ha un periodo di circa 24 ore. Come si può notare dalla parte bassa del pannello a, anche mettendo i topi in condizioni di buio costante, il ritmo circadiano rimane inalterato.
I topi KO per cry1, (pannello b) invece, mostrano un periodo più breve (di 22.5 ore). Questo viene evidenziato solo quando vengono messi in condizioni di buio costante, mentre in presenza di cicli luce/buio normali (12h/12h) si comportano normalmente. Notare come la periodicità rimanga, ne viene solo alterata la lunghezza.
Opposta situazione avviene nei KO per cry2 (pannello c), che invece hanno un periodo più lungo (24.6 ore) e quindi in condizioni di buio completo sono sfasati pian piano “in avanti”. In presenza di singoli KO, tuttavia, una delle due proteine potrebbe compensare, almeno in parte per l’altra.
Quando vennero invece analizzati i doppi KO (che quindi mancano sia di cry1 che di cry2), si notarono due interessanti fenomeni. Innanzitutto i topi avevano ancora una marcata attività circadiana in condizioni di luce/buio (pannello f), indicando che altri sistemi, oltre a cry1/2 possono modulare questi processi. In oltre, in condizioni di buio totale, invece, si ottiene un’immediata perdita di questo ritmo, e l’attività motoria diventa irregolare. I pannelli d ed e, mostrano situazioni intermedie in cui viene mantenuta solo una copia di cry1 o di cry2.
Questi risultati, nel complesso, indicano che il corretto bilanciamento dell’attività di questi due geni è necessario per ottenere un corretto ritmo circadiano.
Infine, a conferma di quanto appena detto, si può osservare che quando si fa passare un topo normale (pannello a) tra due diversi regimi di luce/buio, si nota solo un lento adattamento dell’attività motoria al nuovo “pattern”. I KO per cry1/2, invece, mostrano un immediato cambiamento nell’attività (pannelli c e d).
Molta strada è stata fatta negli ultimi anni per comprendere questi fenomeni, ma c’è ancora molto da studiare e l’argomento è sicuramente affascinante.
Tags:
Animali transgenici,
Neuroscienze,
Ritmi circadiani
21 luglio 2008 - 2:58 pm
Oggi vi parlerò di un tema che trovo particolarmente interessante: la relazione fra ormoni e cervello.
Molti ormoni infatti, oltre ad avere effetti a livello degli organi periferici hanno anche effetti a livello del sistema nervoso centrale. Voglio in particolare soffermarmi su una classe di ormoni, detti “ormoni steroidei”, che sono una serie di ormoni derivati dal colesterolo, dalla struttura chimica molto simile e con moltissime funzioni a livello sia centrale che periferico. Sicuramente tutti voi conoscerete almeno di nome alcuni di questi ormoni, come l’estrogeno, il testosterone, il progesterone ed il cortisolo (il famoso “ormone dello stress“). Tutti questi ormoni hanno effetto a livello del sistema nervoso centrale. La cosa interessante è che gli effetti a livello centrale possono non essere correlati con quelli a livello periferico. Ad esempio, è noto che gli ormoni sopra citati sono coinvolti nel controllo del sistema riproduttivo, della gravidanza etc., ma a livello del sistema nervoso centrale possono avere effetti completamente diversi.
Uno dei primi studi di questi effetti risale all’inizio del ’900, quando venne scoperto che il progesterone (o meglio alcuni suoi metaboliti) hanno effetto anestetico. Nonostante al tempo di questa scoperta non si conoscesse molto a riguardo del funzionamento del cervello, è oggi chiaro che questo effetto deriva dal aumentata sensitività di un tipo di recettori per il GABA (chiamati GABA-A), uno dei maggiori neurotrasmettitori nel sistema nervoso. In presenza di GABA, i recettori GABA-A fanno entrare ioni cloro all’interno della gran parte dei neuroni, portando ad una generale depressione della loro attività. In presenza di progesterone questo effetto del GABA è amplificato e quindi la stessa quantità di neurotrasmettitore porta ad una riduzione più grande dell’attività neuronale, spiegandone quindi l’effetto anestetico.
Altri effetti molto studiati sono quelli dell’estrogeno. E’ noto ad esempio che l’estrogeno può essere neuroprotettivo (ma anche neurotossico a seconda della dose) e favorire la produzione di nuove sinapsi in diverse aree cerebrali, avendo ad esempio effetti sulla memoria.
Insomma, come vedete effetti abbastanza diversi da quelli classicamente noti. Ma la cosa non finisce qui! Il cervello stesso è sede di produzione di quelli che vengono chiamati neurosteroidi, cioè steroidi generati o modificati a livello cerebrale. Le cellule cerebrali (in particolar modo la glia, ma anche i neuroni) possiedono infatti tutto il corredo enzimatico necessario a produrre gli ormoni a partire dal colesterolo, oppure per modificare ormoni prodotti da altri organi. Anche se chimicamente gli ormoni prodotti sono gli stessi, sembra che le funzioni dei neurosteroidi possano essere diverse da quelli degli steroidi periferici. Infatti, la produzione a livello cerebrale permette di concentrare spazialmente e temporalmente la produzione di ormoni, ottenendo così alte concentrazioni in breve tempo, al contrario dei livelli sanguigni di ormoni che sono generalmente molto bassi e cambiano molto lentamente nel corso del tempo.
Quello dei neurosteroidi è un argomento complesso e con aspetti ancora molto controversi. Negli ultimi anni tuttavia, anche grazie al superamento di molte difficoltà tecniche dello studio di questi effetti si stanno cominciando a delineare più chiaramente i meccanismi coinvolti in questi processi.
Infine, a chi fosse interessato all’argomento, consiglio questa bella review di Balthazart e colleghi:
Functional significance of the rapid regulation of brain estrogen action: where do the estrogens come from?
Tags:
Neuroscienze,
Neurotrasmettitori,
Ormoni
17 maggio 2008 - 4:44 am
 Finalmente torno a scrivere sul blog dopo un periodo di latitanza (mi scuso con tutti i lettori, ma sono stato assorbito dalla scrittura della tesi…).
Finalmente torno a scrivere sul blog dopo un periodo di latitanza (mi scuso con tutti i lettori, ma sono stato assorbito dalla scrittura della tesi…).
Oggi vi voglio parlare di un argomento un po’ particolare: la sinestesia. No, non sto parlando della figura retorica…. bensì del fenomeno per cui alcune persone associano determinati stimoli sensoriali associati ad un senso differente.
Il tipo di sinestesia più comune è la sinestesia associata ai grafemi. Un sinesteta associa ad un certo grafema (es. la lettera “A” o il numero “7″) un certo colore. Ad esempio la “A” potrebbe essere associata al rosso: alcune persone vedono effettivamente le lettere colorate, altri creano un’associazione nella loro mente, ma riescono comunque a definire il “vero” colore della lettera. Altri tipi di sinestesia più rari coinvolgono, ad esempio, il gusto: in questo caso la persona percepisce una certa sensazione gustativa quando pronuncia una determinata parola.
Potete leggere un interessante report di un caso di questo tipo in questo articolo (free!)The perceptual reality of synesthetic colors.
L’articolo presenta un’analisi di un caso di sinestesia grafema->colore, in cui il soggetto vede lettere o numeri di un certo colore, anche se sono scritti effettivamente in nero. Uno degli esperimenti consiste nell’identificare un 2 in una figura (generata casualmente dal computer) piena di 5. Il tutto è scritto in bianco su nero, ma il sinesteta ha tempi di risposta più bassi di non-sinesteti a cui viene proposto lo stesso task in quanto per lui il 2 è arancione ed i 5 sono azzurri. Se però la stessa cosa viene fatta con 6 ed 8 che lui vede entrambi in blu, oppure con simboli che non rappresentano una lettera o un numero, i tempi di risposta non sono differenti da quelli dei non-sinesteti.
Un famoso test per individuare sinesteti grafema->colore è l’utilizzo dell’effetto Stroop. Un esempio di questo effetto è il seguente:
dite ad alta voce, ed il più velocemente possibile i colori di queste parole
Rosso Blu Verde Giallo Nero Blu
ora fate lo stesso con queste
Blu Giallo Blu Verde Rosso Rosso Blu
L’effetto Stroop è l’effetto per cui è più difficile dire i colori della seconda serie di parole rispetto alla prima, in quanto nel nostro cervello c’è un’interferenza fra il significato della parola e la percezione della parola stessa. Se ad un sinesteta che vede il numero 6 in rosso viene fatto lo stesso test chiedendogli di dire il colore di questi due numeri:
6 6
la performance nel nominare il colore del secondo sarà più bassa che per il primo.
Il primo report pubblicato di questo fenomeno risale a fine ’800 quando Francis Galton pubblicava su The Journal of the Anthropological Institute of Great Britain and Ireland l’articolo (molto interessante, consiglio di leggerlo) “Visualised numerals“. Le esatte cause di questo fenomeno non sono note, ma sembra che derivi dal fatto che alcune aree della corteccia cerebrale deputate alla percezione di diversi sensi siano collegate in maniera “non corretta“. La sinestesia ha anche una componente genetica ed è infatti una condizione presente in più individui nella stessa famiglia.
Dunque vi chiedo… di che colore è il numero 6?
Tags:
Mente,
Percezione,
Psicologia,
Sinestesia
19 aprile 2008 - 2:02 am
Non c’è dubbio: i ricercatori amano costruire semplici e chiari modelli dei processi biologici che studiano. Tutto sembra così bello quando c’è un schemino corredato di frecce e cascate del segnale che ti chiarifica come vanno veramente le cose… o forse no? Oggi vi faccio un esempio di come a volte la situazione è un po’ più complessa (e meno perfetta) di quanto ci piacerebbe credere.
Parliamo di comunicazione neuronale: il modello classico a riguardo è quello della sinapsi. Citando da Wikipedia: “una sinapsi è una struttura altamente specializzata che consente la comunicazione tra le cellule del tessuto nervoso, i neuroni”. Quello che ciascun libro di testo vi fa vedere è che l’assone di un neurone (il neurone presinaptico) va a contattare un dendrita, il corpo cellulare o l’assone di un altro neurone (il neurone postsinaptico). In questo modo i due neuroni sono connessi e possono parlare fra di loro.
Ma come fanno esattamente? Ci sono due tipi di sinapsi : sinapsi chimiche e sinapsi elettriche. Oggi vi parlerò solo di sinapsi chimiche: non ne farò una descrizione esaustiva, quindi non prendete questo post come testo sacro… non ha nemmeno uno schemino corredato di frecce!
La sinapsi chimica in breve funziona così: il neurone presinaptico viene in qualche modo attivato, non ci interessa come al momento. Questo provoca la generazione di potenziali d’azione (e su dove esattamente questi potenziali d’azione siano generati si potrebbero spendere pagine…) che possono viaggiare sull’assone ed arrivare al terminale, dove abbiamo la nostra sinapsi chimica. Il terminale del neurone è pieno di piccole strutture chiamate vescicole, e piene di una o più determinate sostanze chimiche dette neurotrasmettitori. Quando i potenziali d’azione arrivano al terminale, facilitano l’entrata di calcio nel neurone presinaptico e ciò permette la fusione delle vescicole con la membrana sinaptica ed il rilascio del loro contenuto all’esterno. Il neurotrasmettitore potrà poi legarsi ad opportuni recettori localizzati all’interno della sinapsi sul neurone postsinaptico e attivare o inibire quest ultimo, modulandone la capacità di generare potenziali d’azione e quindi in ultima istanza di rilasciare le sue vescicole di neurotrasmettitore sul neurone successivo. Il neurotrasmettitore viene poi recuperato dai neuroni o dalla glia (l’altra grossa categoria di cellule presente nel cervello) per essere riciclato o alterntativamete distrutto.
Questa immagine, presa da questo sito, mostra una fotografia al microscopio elettronico di una sinapsi. Tutte quelle sferette che vedete sono vescicole sinaptiche che si stanno fondendo con la membrana plasmatica ad una sinapsi.

Bene, quindi è tutto semplice e funziona benissimo… beh non proprio!
Benchè questo fosse un po’ il dogma di come funziona una sinapsi, negli ultimi 15-20 anni sono state scoperti altri modi in cui il nostro cervello utilizza i neurotrasmettitori.
Innanzitutto, partirei dalla glia: la glia è sempre stata considerata come qualcosa di inerte, materiale di supporto per i neuroni. Questo è risultato essere assolutamente falso, la glia ricopre ruoli molto più complessi nel cervello ed uno di questi è proprio quello di produrre neurotrasmettitori! Non solo quindi è implicata nel riciclaggio dei neurotrasmettitori, ma può anche secernerli. Un recente studio ha addirittura mostrato che alcune cellule gliali possono generare potenziali d’azione… non esattamente materiale inerte quindi! Gli stessi trasportatori che permettono di internalizzare il neurotrasmettitore nella glia, possono funzionare al contrario in alcune situazioni ed invece di internalizzarlo lo trasportano all’esterno!
Un’altra modalità di rilascio di neurotrasmettitori è il rilascio non vescicolare: in alcuni casi infatti un po’ di neurotrasmettitore può “scappare” da un neurone senza bisogno che ci sia la fusione di una vescicola con la membrana sinaptica. Questo processo è assolutamente indipendente dai potenziali d’azione e quindi questo neurotrasmettitore viene rilasciato passivamente.
Infine quando il neurotrasmettitore viene rilasciato nello spazio sinaptico, ancora una volta un po’ ne può “scappare via” (neurotransmitter spillage) e andare all’esterno della sinapsi, in sinapsi o all’esterno di sinapsi di altri neuroni. Questo è un bellissimo esempio, a mio parere, di quanto imperfetto sia il nostro cervello, ma di come l’evoluzione ci abbia permesso di sfruttare queste imperfezioni.
Se infatti è vero che questo rilascio involontario di neurotrasmettitore può andare ad attivare altri neuroni che in teoria non avrebbero dovuto partecipare alla trasmissione di quel particolare impulso, è anche vero che il nostro cervello è in grado di sfruttare questa situazione: esistono infatti i cosiddetti recettori extrasinaptici per i neurotrasmettitori, cioè recettori localizzati al di fuori della sinapsi che possono captare queste “perdite” ed attivare segnali intracellulari. Se pensate che un metodo di comunicazione puramente basato sulla casuale perdita di una sostanza dalla sinapsi sia inefficiente e poco “smart”… beh, vi do ragione, ma sappiate che esistono cellule nel cervelletto che comunicano esclusivamente utilizzando perdite di glutamato da sinapsi vicine!
Tags:
Attività neuronale,
Firing,
Neuroscienze,
Potenziali d'azione
11 aprile 2008 - 12:47 pm
Come abbiamo già detto in passati post i neuroni comunicano tra loro grazie alla generazione di “scariche elettriche”, chiamate potenziali d’azione. E’ quindi interessante avere la possibilità di generare artificialmente dei potenziali d’azione per studiare le proprietà di un certo neurone o di un network di neuroni. Ad esempio uno può stimolare il neurone 1 e vedere cosa succede al neurone 2 che vi è collegato.
Il classico approccio per fare ciò è utilizzare un elettrodo (un piccolo filamento di argento) inserito in una micropipetta di vetro dalla punta di pochi millesimi di millimetro di diametro e riempita di una soluzione conduttrice. Questo microelettrodo viene messo a contatto con la cellula (rompendone o meno la membrana a seconda del tipo di esperimento) e permette di iniettare con precisione corrente nella cellula, oltre a poterne rilevare l’attività elettrica spontanea.
Esistono però diversi altri approcci per eccitare o inibire un neurone che non richiedono l’utilizzo di un microelettrodo. I vantaggi di non usare microelettrodi sono molteplici, ma i principali sono: 1) andare a piazzare la minuscola punta dell’elettrodo sulla membrana cellulare non è proprio la cosa più semplice del mondo… 2) non si va a disturbare meccanicamente la cellula 3) sono più alla moda oggigiorno, tanto che qualcuno ne parla in un blog 
Uno di questi metodi è quello di utilizzare un neurotrasmettitore come, ad esempio, il glutamato. Possiamo “spruzzare” una piccola quantità di glutamato sulla cellula e questa, nella maggior parte dei casi, verrà eccitata. Esistono molte variazioni sul tema, ma oggi vi parlerò di un nuovo lavoro, apparso su Nature Methods di questo mese, che ha introdotto un nuovo interessante approccio.
Il lavoro in questione è il seguente: Photochemical control of endogenous ion channels and cellular excitability. – Fortin et al. – Nat. Methods 2008 Apr;5(4):331-8.
Gli autori hanno generato un “photoswitchable affinity label” (PAL), una piccola molecola che si lega selettivamente a canali sulla membrana del neurone che fanno passare gli ioni potassio (K+). In particolare, questa molecola lega un tipo di canali per il potassio (voltage-gated potassium channels) che sono sensibili a variazioni di voltaggio nella cellula. La normale funzione di questi canali è quella di riportare la cellula allo stato basale dopo la generazione di un potenziale d’azione. Insomma hanno un effetto “calmante” sull’attività cellulare.
PAL può legarsi selettivamente a questi canali e bloccarli, impedendo così il passaggio di potassio, risultando quindi in un eccitazione della cellula. La cosa interessante è che PAL è “photoswitchable”, cioè può essere “accesa” o “spenta” utilizzando luce di colori differenti. Questo è dovuto al fatto che PAL può cambiare forma quando viene colpita da luce di un certo colore: utilizzando luce viola PAL non blocca il canale, utilizzando luce verde lo blocca.
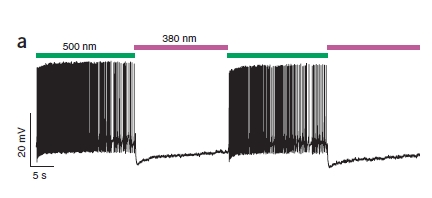
Il risultato è facilmente visualizzabile in questa figura:
Come vedete, quando la cellula è colpita da luce verde è iperattiva (ogni linea verticale rappresenta un potenziale d’azione, in alcune parti sono così fitti da non poterli distinguere) perchè PAL sta bloccando i canali al K+, mentre la luce viola silenzia la cellula.
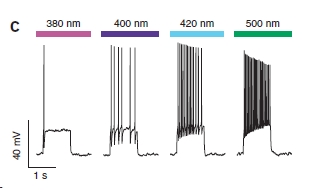
E non è finita così! Il funzionamento è graduale, quindi utilizzando luce di colore intermedio si ottiene una parziale eccitazione della cellula.
Certo, tutto questo è stato fatto su cellule in coltura ma sarebbe molto interessante vedere questo tipo di tecniche utilizzate in vivo (non dubito che ci si arriverà presto…). Ovviamente lo stesso principio potrebbe essere applicato a qualsiasi altro canale sui neuroni, si tratterà solo di sintetizzare molecole specifiche che permettano di legarsi ad altri canali!
Tags:
Attività neuronale,
Canali voltaggio dipendenti,
Elettrofisiologia,
Firing,
Imaging,
Neuroscienze,
Potenziali d'azione
14 marzo 2008 - 10:59 pm
(…continua dal post precedente)
Come dicevamo nella prima parte di questo post i neuroni mirror giocano un ruolo importante nell’imitazione. In questa seconda parte vi mostrerò un esempio pratico del funzionamento di questi neuroni, parlandovi di un articolo apparso sull’ultimo numero di Nature:
Precise auditory-vocal mirroring in neurons for learned vocal communication – Nature 2008 Jan 17;451(7176):305-10.
Questo articolo mostra la presenza di neuroni mirror nel cervello del passero che vengono attivati quando l’uccello canta una sua canzone e quando sente la stessa canzone cantata da un altro uccello (o sente una registrazione della sua canzone).
Gli uccelli canori, infatti, sono noti per imitare il verso di altri uccelli e i piccoli di queste specie imparano a cantare imitando i propri genitori.
Per effettuare questi esperimenti Prather e colleghi hanno sfruttato il fatto che molti uccelli canori hanno una tendenza al controcanto: quando sentono un altro uccello cantare nel loro territorio, infatti, rispondono cantando anche essi. Questo può essere un gesto di “sfida” nei confronti di un uccello di un’altra specie o ad esempio un gesto di riconoscimento di un “familiare”.
I ricercatori hanno quindi utilizzato registrazioni del canto di vari uccelli e sono riusciti a trovare dei mirror neurons che sono attivati sia quando l’uccello sente la registrazione sia quando vi risponde.

Questa immagine fa vedere un esempio della risposta di neuroni auditori. Nella prima colonna è mostrata la risposta alla canzone primaria dell’uccello. La prima traccia in alto rappresenta l’attività di un singolo neurone, e ciascuno dei picchi che vanno in basso è corrispondente ad un potenziale d’azione, indice di attività di quel neurone. L’attività è massima in corrispondenza dello stimolo auditorio (che vedete nell’ultima riga).
Se confrontate questa risposta con la risposta ad un altra canzone dello stesso uccello (colonna 2) o di un altro uccello (colonna 3) potete notare come in questi ultimi due casi non ci sia corrispondenza fra lo stimolo e i potenziali d’azione.
La cosa è ancora più chiara guardando la seconda riga che mostra la risposta del neurone a diverse presentazioni dello stimolo. Nel grafico ciascun puntino rappresenta un potenziale d’azione e ciascuna riga rappresenta un diverso trial su quel neurone. Come potete vedere la risposta è quindi altamente riproducibile.
Ma cosa succede durante il controcanto? Beh, ecco un esempio dei risultati ottenuti:

In a) l’uccello è esposto ad una registrazione della sua canzone primaria, e il neurone è attivo durante quel periodo. A questo punto l’uccello esegue un controcanto in risposta (con la stessa canzone) e il neurone è ancora attivo. Se però la canzone ascoltata e quella cantata differiscono (b e c) il neurone è attivo solo in una delle due situazioni! Insomma, una cosa molto simile a quanto visto nelle scimmie da Rizzolatti e colleghi (vedi post precedente).
Infine gli autori mostrano anche che queste risposte sono specifiche per le singole note del canto. Alcuni neuroni rispondono durante una particolare nota in una sequenza: facendo ascoltare la sequenza al contrario la risposta è molto attenuata, ma invece persiste se si fa ascoltare una registrazione della canzone di un altro uccello che abbia note simili.
Concludendo, questo studio mostra un altro interessante esempio di come il cervello interpreti con precisione le informazioni derivate dall’ambiente e di come l’imitazione sia codificata a livello neuronale. Situazioni simili a quella descritta in questo articolo potrebbero anche essere alla base dell’apprendimento del linguaggio nell’uomo, probabilmente integrate anche da altri stimoli (visuali e motori).
Tags:
Elettrofisiologia,
Imitazione,
Percezione,
Potenziali d'azione
4 marzo 2008 - 2:06 am
A tutti noi è capitato di dover imparare a fare qualcosa: come è noto uno dei modi migliori è iniziare guardando qualcun altro che fa ciò che vogliamo imparare. Che si tratti di suonare uno strumento, usare un macchinario in laboratorio o imparare a parlare, infatti, l’imitazione gioca sicuramente un ruolo importante nel nostro processo di apprendimento. Ma come funziona esattamente tutto ciò? Abbiamo già detto in post precedenti (ad es. questo e questo) che il nostro cervello memorizza ed impara attraverso il rafforzamento di alcune sinapsi e l’indebolimento di altre… ma come entra l’imitazione in questo sistema?
Purtroppo non vi posso dare una risposta completa, ma vi posso raccontare almeno parte della storia.

Un macaco neonato imita un ricercatore che tira fuori la lingua!
(da Evolution of Neonatal Imitation. Gross L, PLoS Biology Vol. 4/9/2006, e311)
Tutto cominciò circa una decina di anni fa con gli studi di Giacomo Rizzolatti e colleghi all’università di Parma, i quali stavano studiando l’attività dei neuroni della corteccia premotoria del macaco, una regione coinvolta nella pianificazione delle azioni e nella decisione di quali atti compiere (da cui il nome premotoria). Ad esempio, alcuni neuroni di questa regione potrebbero venire attivati quando il macaco prende un pezzo di cibo da un piatto per metterlo in bocca, altri potrebbero essere attivati quando invece si arrampica su di un albero.
Durante i loro studi, Rizzolatti e colleghi scoprirono l’esistenza di una sottopopolazione (10-20%) di questi neuroni, i quali vengono attivati sia quando l’animale fa una certa azione (es. prende una banana), sia quando vede un altro animale fare la stessa azione. Questi neuroni furono chiamati mirror neurons o neuroni specchio e sembrano essere degli ottimi candidati per spiegare questi processi di apprendimento per imitazione.
La precisione di questi neuroni è notevole: ad esempio, un certo neurone che veniva attivato quando la scimmia prendeva il cibo, veniva anche attivato quando vedeva il ricercatore prendere il cibo. Se però quest ultimo usava delle pinze per prendere il cibo l’attivazione era molto minore, e praticamente nulla se faceva il gesto di prendere qualcosa, ma senza che effettivamente ci fosse del cibo.
Da allora, molti studi sono stati fatti nel campo dei neuroni mirror che sono stati trovati anche nell’uomo e in alcune specie di uccelli (come vedremo nella seconda parte di questo post).
In particolare sembra che questi neuroni siano molto importanti nei processi di apprendimento del linguaggio e altri studi hanno suggerito che un loro malfunzionamento potrebbe essere in parte implicato nell’autismo.
Per chi fosse interessato, ecco il link ad uno degli articoli di Rizzolatti: Action recognition in the premotor cortex
(continua…)
Tags:
Imitazione,
Memoria,
Mente,
Neuroscienze,
Percezione,
Plasticità sinaptica,
Visione


